
- Title
-
Differential requirements for myogenic regulatory factors distinguish medial and lateral somitic, cranial and fin muscle fibre populations
- Authors
- Hinits, Y., Osborn, D.P., and Hughes, S.M.
- Source
- Full text @ Development

Myf5 or Myod are required for slow fibre formation. Dorsal flatmounts of zebrafish embryos injected with the indicated MOs and uninjected controls (con), analysed at the 13- to 15-somite stage by whole-mount immunohistochemistry for MyHC (red) or in situ mRNA hybridisation for the indicated mRNAs (blue). Anterior to top. (Left panels) myf5+myod MOs ablated all slow myosin heavy chain (smyhc1 and slow MyHC, arrowheads), but did not completely prevent actin or tpma expression (arrowheads). myod MO slightly delayed smyhc mRNA and MyHC accumulation (arrows), whereas other single MOs had little effect. (Right panels) Single MRF MOs had no effect on adaxial expression of the other MRFs. However, the myod MO reduced myog signal in fast muscle precursors (white arrows). The box region is shown magnified beneath. Note the increase in signal for each MRF caused by its cognate MO (asterisks). s, somites. EXPRESSION / LABELING:
|

myf5hu2022 mutants appear normal as embryos but fail to mature into adults. (A) Myf5 truncation predicted from the myf5hu2022 allele. The location of the introduced stop codon is shown; B, basic domain; HLH, helix-loop-helix domain. (B) Genotyped zebrafish embryos from a myf5+/hu2022 carrier incross show no phenotype at 5 days post-fertilisation (5d) as compared with the wild type (+/+). (C) Survival at various ages of fish of each genoptype obtained from myf5+/hu2022 incrosses. The numbers of embryos are shown. (D) In situ mRNA hybridisation for myf5 revealed mildly reduced signal in putative myf5+/hu2022 (13/28) and lower levels in putative myf5hu2022/hu2022 (9/28) embryos. No such effect was observed in myf5 morphants (0/27). (E) Embryos from an myf5+/hu2022 incross lack adaxial myog mRNA early (15/49), and later have a mild delay in myog mRNA accumulation in fast muscle precursors correlated with reduced myf5 mRNA (20/74). Scale bars: 100 μm in D; 150 μm in E. EXPRESSION / LABELING:
|

Myf5 requirement differs between somitic, head and fin muscle development. In situ mRNA hybridisation and/or MyHC immunohistochemistry of control and morphant wild-type or myf5+/hu2022 incross embryos. Dorsal flatmounts with anterior to top (A,F), or whole-mounts with anterior to left in ventral (B, upper row, C,D), lateral (B, lower row) or dorsal (E) view. (A) myf5hu2022 mutants injected with myod MO show a loss of the adaxial slow muscle differentiation markers smyhc1, actin, tpma and mef2ca and MyHC immunostaining (black arrows), but retain other adaxial markers such as prdm1 and ptc1 (white arrows). (B-D) Head muscles are present in 80 hpf myf5hu2022 mutants but are missing from embryos injected with myod (B,C) or myog MO (C). Arrowheads indicate lack of ventral (green), dorsal (yellow), extra-ocular (red) and sternohyoideus (sh, white) muscles. All embryos show normal heart (h, blue arrow). Note the presence of sternohyoideus in myog morphants. All embryos from a myf5+/hu2022 incross have normal expression of myhz1 at 72 hpf (C) and myog at 48 hpf (D) in the head. myod morphants lack myog mRNA (19/21), although sternohyoideus was still detected in some embryos (6/21). (E) In pectoral fin, myog mRNA in muscle masses (pink arrowheads) and MyHC are ablated or reduced by loss of Myod, but not by loss of Myf5. Ibx2 mRNA is unaffected by myod MO. (F) Expression of hsp90a is delayed in fast muscle precursors (arrow) of the myf5hu2022 mutant and recovers in somites (arrowhead), but is drastically reduced throughout the axis in myf5hu2022 mutants injected with myod MO, both in slow and fast precursors. meox1 expression is unchanged in fast precursors. EXPRESSION / LABELING:
PHENOTYPE:
|

Myf5 or Myod is required for medial fast myogenesis. In situ hybridisation for myog, myhz1, tpma, mylz2 or mef2d or immunohistochemistry for the indicated myosins in wild-type or myf5+/hu2022 incross zebrafish injected with the indicated MOs. Lateral views, anterior to left unless otherwise stated. (A) myog expression was delayed in embryos injected with myf5+myod MOs. Dorsolateral views, anterior to top. (B) MyHC was reduced in both epaxial and hypaxial domains in myod MO embryos, grossly reduced in double myf5+myod morphants and further diminished in myf5hu2022 mutants injected with myod MO. Note the complete absence of residual cells in some individual somites (dotted outline). (C) Residual differentiated muscle in myf5+myod morphants expressed fast but not slow MyHC at 24 hpf. Lateral view of confocal stack of somites 9-10. (D) Fast muscle markers were indistinguishable in 24 hpf myf5hu2022 homozygous mutants and their siblings (left column), reduced by myod MO in siblings (middle) and almost eliminated by myod MO in myf5hu2022 mutants (right). Inset illustrates that myf5 MO does not worsen the myod MO plus myf5hu2022 phenotype. mylz2 mRNA accumulation (red, arrowheads) and myog downregulation (blue) are delayed at ∼20 ss in fast muscle. (Bottom panels) Transverse sections of double knockdowns show residual fast muscle (arrowheads) immediately lateral to adaxial and/or twist2-expressing sclerotomal cells (arrows). (E) (Left, top) mef2d mRNA initiates normally in pbcs in the myod morphant, but is restricted to the medial region of rostral somites (yellow brackets). In myf5+myod MOs, mef2d mRNA is absent. Dorsal flatmounts, anterior to top. (Left, bottom) Lack of both myf5 and myod ablates mef2d expression. Lateral views, anterior to top. (Right) By 22 ss, transverse sections (at the level of the bar on lateral flatmounts) show reduction of mef2d mRNA (purple) in myod or myog, but not in myf5, single morphants. Mef2 position is shown relative to slow fibres (brown). Note the expression of mef2d lateral to migrating slow fibres in the control and in myf5 and myog, but not myod, morphants (arrows). EXPRESSION / LABELING:
|
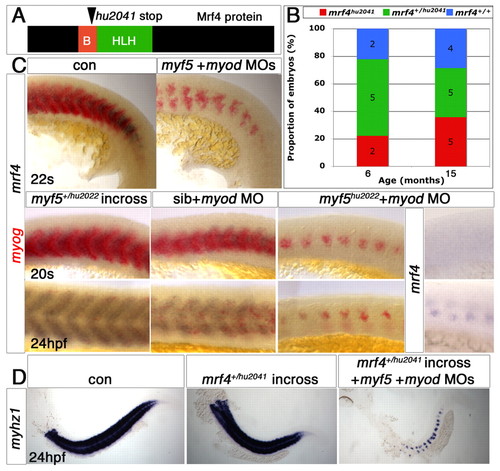
Mrf4 has no role in trunk muscle differentiation. (A) Schematic illustrating the location of the mutation encoded by mrf4hu2041. (B) Proportion of mrf4hu2041/hu2041 (red), mrf4hu2041/+ (green) or mrf4+/+ (blue) in progeny from heterozygote crosses. The number of embryos of each genotype is shown. Homozygote mutants survive well and without obvious defects. (C) Whole-mount in situ mRNA hybridisation of myf5+myod morphants or myod MO-injected myf5hu2022 mutants reveal that residual medial fast fibres express myog (red) but not mrf4 (purple) at 22 ss, but accumulate mrf4 mRNA after terminal differentiation at 24 hpf. Lateral view, anterior to left. (D) In situ mRNA hybridisation for myhz1 (purple) in embryos from a mrf4+/hu2041 incross reveals that injection of myf5+myod MOs fails to ablate all muscle irrespective of mrf4 genotype. EXPRESSION / LABELING:
|

Loss of Myod and Myog ablates somitic fast muscle. In situ mRNA hybridisation (A,B,E) or immunofluorescent confocal stacks (C,D) of 24 hpf wild-type (con), Tg(acta1:GFP) or myf5+/hu2022 incrosses injected with the indicated MOs. Lateral views, anterior to left. (A) Magnified regions (boxed) showing that lack of Myf5, Myod and Myog results in loss of all muscle. Knockdown of Myod and Myog results in loss of most fast, but not slow, muscle. Note that mylz2 (red) is weakly expressed in immature slow muscle. (B) Loss of Myog and Myf5 has little effect, if any, on fast myhz1 or slow smyhc1 expression. (C) Injection of myod+myog MOs into Tg(acta1:GFP) fish results in loss of fast muscle, as shown by loss of Mef2 staining and GFP medial to the slow fibre layer accompanied by somitic apoptosis as revealed with DAPI (blue in the optical transverse sections). Elongated mononucleate slow fibres differentiate and express GFP, slow MyHC and Mef2. myod morphants confirm the lateral reduction of fast muscle mass. (D) myod+myog MOs ablate fast MyHC in somites, whereas single MOs have a lesser effect. (E) Titration of myod+myog MOs progressively diminishes residual myhz1-expressing cells. The lower panels are high-magnification images of mid-trunk somites (boxed). |

Residual fast fibres after Myf5 plus Myod knockdown are hedgehog dependent. In situ mRNA hybridisation (A,B) and immunofluorescent confocal stacks (C) of 23 ss, 24-26 hpf wild-type, Tg(mylz2:GFP) or myf5+/hu2022 incross embryos. Lateral views, anterior to left. (A) Cyclopamine (CyA) treatment of embryos from a myf5+/hu2022 incross injected with myod MO ablates residual myhz1 expression (arrows, magnified in insets). Fast muscle still forms in the absence of Myod and Hh signals. Note the altered somite shape, paralleling the absence of slow fibres. (B) Expression of eng1a and eng2a (arrowheads) is missing from myf5hu2022 mutants injected with myod MO and is reduced in myod+myog double morphants. Eng1a is not affected by myf5 mutation (revealed by weak myf5 mRNA expression) or myod MO alone, although eng2a is drastically reduced in the myod morphant. (C) Tg(mylz2:GFP) embryos injected with myod MO show reduction of both the weak Engrailed protein in MFF nuclei (yellow arrowheads) and the strong Engrailed expression of muscle pioneers (blue arrowheads). (D) Schematic illustrating how the cells generated in medial and lateral modes of myogenesis in the early zebrafish somite (left) each contribute to several later compartments (right). Note that dermomyotomal contributions are not defined because cell lineage is largely unknown. EXPRESSION / LABELING:
|
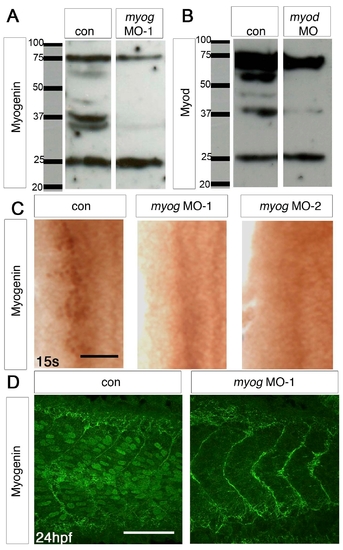
myog and myod MOs can specifically block translation of their mRNAs. (A,B) Western blot of protein extracts from control (A,B) and myog MO-1 (A) or myod MO (B), showing absence of specific bands at ∼37 kDa from the MO lanes. Myod immunohistochemical stain is also ablated (Hammond et al., 2007) #8381. (C) Immunohistochemistry with Myogenin antibody (brown nuclei) of myog MO-1-injected, myog MO-2-injected and control 15 ss embryos. Dorsal flatmounts of one side of anterior somites, anterior to top. (D) Confocal stacks of tail somites of 24 hpf control and myog MO-1-injected embryos after immunodetection of Myogenin (green nuclei). Lateral view, anterior to left. myog morphants lack the nuclear Myogenin reactivity of control embryos. |
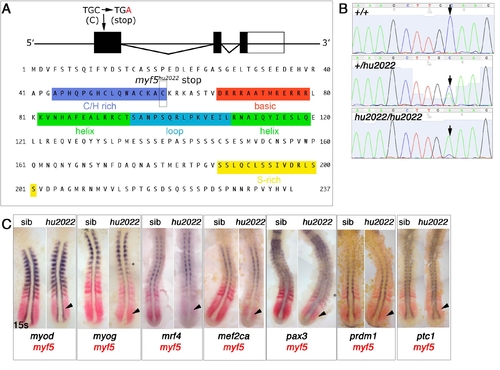
myf5hu2022 mutation generates a likely null allele. (A) The genomic and protein structure of myf5 showing the nature of the hu2022 allele. (B) DNA sequencing traces from three individuals of each genotype. (C) Double in situ mRNA hybridization of myf5 (red) and the gene indicated (purple) in myf5+/hu2022 incross embryos at 13-15 ss. Dorsal flatmounts, anterior to top. myf5hu2022 mutants have low myf5 signal (arrowheads) compared with their siblings, but show no other change in mRNA expression. |
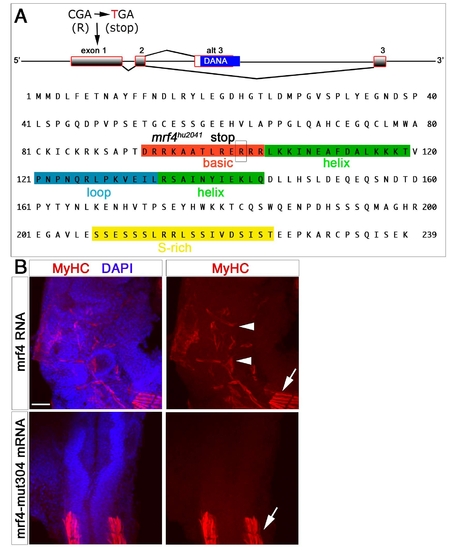
mrf4hu2041 mutation generates a likely null allele. (A) The hu2041 mutation in mrf4 and the resultant truncated protein. (B) Ectopic myogenesis in the head is induced by injection of wild-type mrf4 mRNA, but not by mrf4hu2041 mutant mRNA. The full-length coding sequences of mrf4 and mrf4hu2041 were inserted into the SacI-SalI sites of pβUT3 (gift from Dr Adam Rodaway, KCL). Capped mRNAs were transcribed from linearised plasmid using the T3 mMESSAGE mMACHINE Kit (Ambion), 100 pg/embryo injected at the 1- to 2-cell stage and embryos analysed by MyHC immunodetection at 24 hpf. |