
- Title
-
Chordin expression, mediated by Nodal and FGF signaling, is restricted by redundant function of two beta-catenins in the zebrafish embryo
- Authors
- Varga, M., Maegawa, S., Bellipanni, G., and Weinberg, E.S.
- Source
- Full text @ Mech. Dev.
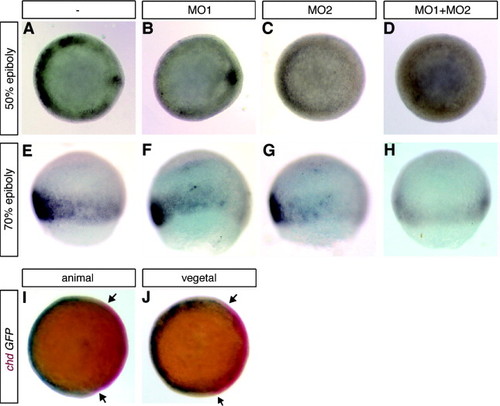
The ventrolateral domain of Wnt/β-catenin signaling is dependent on both β-catenin-1 and β-catenin-2, whereas organizer β-catenin-mediated gene activation is dependent only on β-catenin-2. (A–H) TOP-GFP embryos were injected with βcat1MO (MO1) (B and F), MO2 (C and G), or both βcat1MO and βcat2MO (MO2) (D and H) and compared to uninjected embryos (A and B) after in situ hybridization with probe for GFP at 50% epiboly (A–D) or 70% epiboly (E–H). Embryos are shown in animal view (A–D) or lateral view (E–H). (I and J) chd expression is excluded from the ventrolateral Wnt/β-catenin signaling domain. TOP-GFP embryos at 70% epiboly were assayed by two color in situ hybridization for both GFP (blue) and chd (red) expression. A single embryo is presented in both animal pole (I) and vegetal pole (J) views to show the complementarity of chd and Wnt signaling domains at this stage. The arrows in (I and J) point to a slight region of overlap of the chordin expression and Wnt signaling domains. All embryos are shown with dorsal to the right; (E–H) animal pole is to the top. |
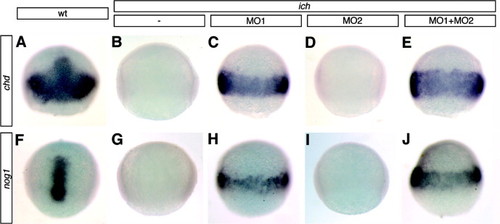
Elimination of β-catenin signaling induces the ectopic expression of dorsal genes around the germ-ring. At 75% epiboly, wild-type embryos have characteristic patterns of chd (A) and nog1 (F) expression, whereas ich embryos do not express these genes (B and G). Injection of βcat1MO (MO1) into ich embryos induces expression of chd (H) and nog1 (H) around the germ-ring, but βcat2MO (MO2) has no effect (D and I). Co-injection of the two MOs into ich embryos causes somewhat increased expression of chd (E) and nog1 (J) around the germ-ring. Concentrations of MOs injected were 3 mM for the single MO1 or MO2 injections and 3 mM each for the double MO injection. Wild-type embryos are shown in dorsal view (A and F) and all other embryos are shown in lateral views. Expression (or lack of) was identical for all embryos for each treatment (>16 in each case) except for (I) in which 1/19 embryos showed very weak expression. EXPRESSION / LABELING:
|
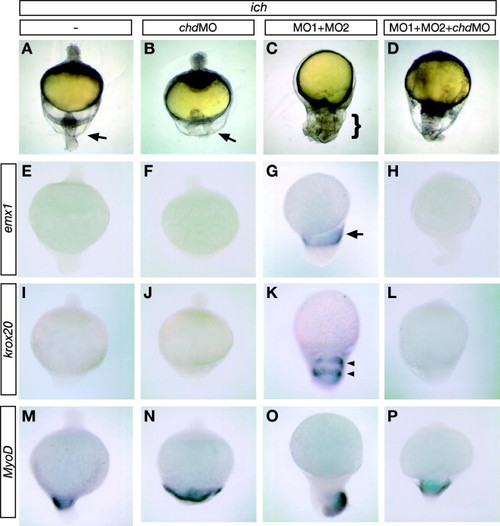
chordin expression is required for expression of neural markers when both β-catenins are inhibited. 24 hpf untreated ich embryos (A, E, I, and M), and ich embryos injected with chdMO alone (B, F, J, and N), βcat1MO and βcat2MO ([MO1 + MO2] C, G, K, and O), or all three MOs (D, H, L, and P) were photographed alive (A–D) or after in situ hybridization with probe for emx1 (E–H) krox20 (I–L), or myoD (M–P). ich embryos, which are highly ventralized if untreated (A), do not express emx1 (E), or krox20 (I), but do express myoD in the tail region (M). When depleted of β-catenin-1 and β-catenin-2 expression by MO treatment, embryos develop the ‘ciuffo’ phenotype (C, bracket indicates the protrusion that has very different morphology from the tail region of ich embryos, indicated by arrow in A) and express emx1 (G, arrow points to single band of expression), and krox20 (K, arrowheads point to two bands of expression), in addition to myoD (O). ich embryos injected with chdMO alone remain highly ventralized, with a broader but less protrusive tail (B, arrow) than in untreated ich embryos, and show no induction of emx1 (F) or krox20 (J). When ich embryos are injected with chdMO as well as the two β-catenin MOs, the posterior protrusion is no longer ‘ciuffo-like’, but rather is tail-like, and the anterior necrotic protrusion (at the end of the embryo opposite the arrow) seen in ich embryos (A) and embryos treated with chdMO alone (B) fails to form (D). The embryos treated with the three MOs are inhibited in both emx1 (H) and krox20 (L) expression but do express myoD in the tail region (P), indicating that the failure of emx1 and krox20 expression in the triple MO-injected embryos was not due to non-specific toxic effects of the MOs. Over 24 embryos were examined for each marker and each condition, with results identical to embryo shown in the panel (except for panel N in which 1 of 24 embryos did not express myoD and panel K in which 2 of 28 embryos did not express krox20). Concentrations of MOs injected were 3 mM for each of the β-catenin MOs and 0.6 mM chdMO. |
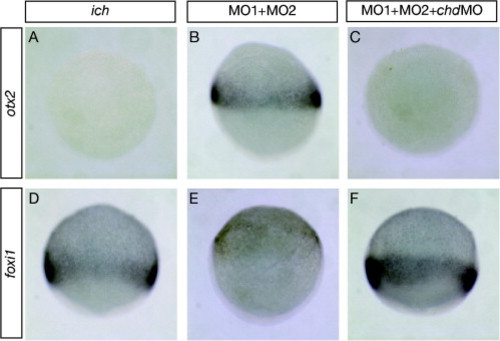
chordin expression is required for otx2 expression and suppresses foxi1 expression in 10 hpf embryos inhibited in expression of both β-catenins. Untreated ich embryos (A and D) and ich embryos injected with βcat1MO and βcat2MO ([MO1 + MO2] (B and E), or additionally with chdMO (C and F) were assayed for expression of otx2 (A–C) or foxi1 (D–F). Inhibition of expression of the two β-catenins results in induction of the anterior neural marker otx2 at the expense of expression of the epidermal marker foxi1. The inhibition of Chd in such embryos prevents otx2 expression and restores the foxi1 expression. Concentrations of MOs injected were as in Fig. 3 and embryos are shown in lateral views. EXPRESSION / LABELING:
|
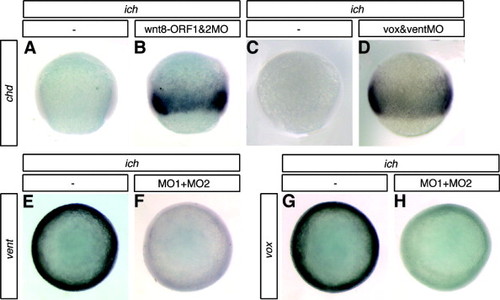
Germ-ring expression of chordin is repressed by Wnt8 through the transcriptional repressors Vox and Vent. ich embryos co-injected with MOs targeted against both ORFs of wnt8 (B) or with MOs against both vox and vent (D) show ectopic expression of chd around the germ-ring. Uninjected ich embryos do not express chd (A and C). Suppression of β-catenin signaling in the germ-ring of ich embryos by injection of βcat1MO + βcat2MO (MO1 + MO2) causes a decrease of vox and vent expression (compare E with F, and G with H). Embryos are shown in lateral (A–D) or animal (E–H) views at 70% epiboly. Concentrations of MOs injected were 0.6 mM of each of the two Wnt8MOs, 0.7 mM of each of voxMO and ventMO, and 3 mM of each of the two β-catenin MOs. EXPRESSION / LABELING:
|
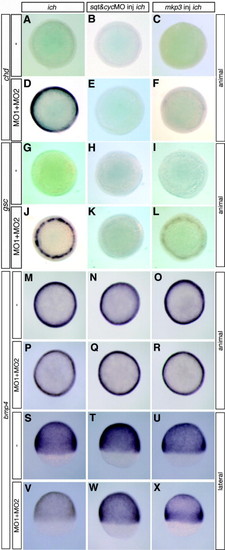
Germ-ring expression of chordin and goosecoid in embryos with impaired Wnt8/β-catenin signaling is dependent on Nodal and FGF signaling pathways. Circumferential germ-line expression of chd (D) and gsc (J) in ich embryos depleted of β-catenins by co-injection of βcat1MO and βcat2MO (MO1 + MO2) was eliminated in embryos co-injected with additional MOs against sqt and cyc (E and K) or with mkp3 RNA to inhibit FGF signaling (F and L). Uninjected control ich embryos and ich embryos injected solely with sqtMO + cycMO or with mkp3 RNA did not express chd (A–C) or gsc (G–I). The widespread ubiquitous expression of bmp4 in ich embryos (M and S) is somewhat reduced when the embryos are co-injected with βcat1MO and βcat2MO (P and V). Injection of ich embryos with sqtMO + cycMO (N and T) or with mkp3 RNA (O and U) does not reduce bmp4 expression. Injection of the four MOs, sqtMO + cycMO + βcat1MO + βcat2MO (Q and W), or mkp3 RNA + the two βcat MOs (R and X), also did not reduce bmp4 expression, except in the latter case, the expression was less uniform with greater expression towards the margin. Results for eve1 expression were essentially identical to that of bmp4 (data not shown). Although only one embryo is shown per panel, results were completely uniform for all embryos in each panel (n > 25 in each case). Embryos are all at 70% epiboly and are shown in animal views (A–R) and for the embryos of (M–R), also shown in lateral view (S–X). EXPRESSION / LABELING:
|
Reprinted from Mechanisms of Development, 124(9-10), Varga, M., Maegawa, S., Bellipanni, G., and Weinberg, E.S., Chordin expression, mediated by Nodal and FGF signaling, is restricted by redundant function of two beta-catenins in the zebrafish embryo, 775-791, Copyright (2007) with permission from Elsevier. Full text @ Mech. Dev.