
- Title
-
The zebrafish maternal-effect gene mission impossible encodes the DEAH-box helicase Dhx16 and is essential for the expression of downstream endodermal genes
- Authors
- Putiri, E., and Pelegri, F.
- Source
- Full text @ Dev. Biol.

Gastrulation defects in maternally mutant mis embryos. Wild-type embryos (A,C) and mis mutant embryos (B,D). mis embryos exhibit reduced epiboly of the blastoderm, as evidenced by the increased thickness of the blastoderm at the animal pole (white solid lines) and reduced coverage of the yolk (white dotted line indicates blastoderm margin). The extent of involution of the hypoblast is also reduced (black brackets). Lateral views (animal top, dorsal right). (E–I) Defects in epiboly and convergence. (E) Labeling strategy, by uncaging a small group of cells at the lateral margin in wild-type (F) and mutant (G) embryos (SV: side view; DV: dorsal view). At a time corresponding to 90% epiboly in the wild-type, the extension of labeled cells along the animal–vegetal axis (double-headed arrows) is reduced in the mutant (I) compared to wild-type (H). In addition, the width of the band of labeled cells (brackets), indicative of cell intercalation during convergence, is reduced in mutant embryos. (F,G) are side views, dorsal to the right. (H,I) are centered on the labeled cells, corresponding to the dorsal view (DV) in (E). Uncaging was carried out at 5:30 h.p.f. and time after fertilization is indicated in h:min. Embryos shown in (F–I) are representative of 5 wild-type and 5 mutant embryos. |

Defects in cell movements in maternally mutant mis embryos. (A–E) Defect in cell mixing during early epiboly. (A) Labeling strategy, by uncaging fluorescent tracer at the sphere stage. Uncaging of a group of cells at the sphere stage in wild-type (B) and mis mutant (C) embryos is followed by extensive cell mixing in the wild-type (D) but little mixing in the mutant (E). (B–E) are side views, tilted toward the animal pole. Time after fertilization as indicated in h:min. Uncaging was carried out at 4:30 h.p.f., and some cell mixing has already occurred in (B,C). Embryos shown in (A–E) are representative of 5 wild-type and 5 mutant embryos. (F–X) Cell autonomy of movement defect. (F) Labeling strategy, by co-transplantation of rhodamine-labeled cells from wild-type (red) and fluorescein-labeled cells from maternally mutant mis (green) donors at the sphere stage (4 h.p.f.) into isochronic unlabeled wild-type host embryos. (G–L) Wild-type cells visualized with the rhodamine channel. (M–R) Mutant cells visualized with the fluorescein channel. (S–X) Merged images. Chimeric embryos were monitored for the first 24 h of development, and a subset of time points is shown (h:min p.f., indicated in (S–X)). Wild-type cells populate the embryonic axis of the host, while mutant cells remain in regions overlying the yolk cell. The embryo shown in (G–X) is representative of transplantations into 30 donor embryos. All panels are lateral views with dorsal to the right, except (L,R,X) with dorsal at the top and anterior to the left. PHENOTYPE:
|

Induction of endoderm-specific genes is affected in maternally mutant mis embryos. Analysis of germ layer gene markers in wild-type (A,C,E,G,I,K,M) and maternally mutant mis embryos (B,D,F,H,J,L,N). Ectodermal markers chordin (A,B) and foxb1.2 (C,D) are induced in wild-type and mis mutants at similar levels, although their anterior-most domains of expression (arrows in A,C) are absent in mis mutants. Mesodermal marker genes no tail (E,F), snail1a (G,H), and lhx1a (I,J) are also induced in the blastoderm margin at normal levels in mis mutants compared to wild-type. On the other hand, mis mutant embryos fail to activate endodermal markers foxA2 (K,L) and sox17 (M,N) in the “salt and pepper” pattern characteristic of endodermal cells (brackets in K,M, compare to no detectable expression in L,N). The axial mesodermal domain of foxA2 (arrowheads in K,L) does become induced in mutant embryos, albeit in an abnormally shaped domain likely caused by the morphogenesis defects associated with the mutation (a similar effect is observed with regards to the axial domain of zlim-1 arrowhead in (I); see also Supp. Fig. 2). Expression of sox17 in forerunner cells (arrowhead in M) is absent in the mutant (N) (see text). Stages are 5 h.p.f. (E,F), 6 h.p.f. (G–J,M,N), and 8 h.p.f. (A–D,K,L). All images are dorsal views with the animal pole at the top, except (E,F) which are animal views. EXPRESSION / LABELING:
|

Defects in endoderm-specific gene expression in maternally mutant mis embryos occur in downstream steps within the pathway. Wild-type (A,C,E,G,I,K) and mis mutant (B,D,F,H,J,L) embryos assayed for expressed transcripts encoding endoderm signaling components, including the nodal signals squint (A,B) and cyclops (C,D), and the transcription factors bon/mixer (E,F) and casanova/sox32 (cas, G-L). Expression patterns for squint, cyclops, and bon appear similar in wild-type and mis mutant embryos. cas becomes induced in the YSL at 5 h.p.f. in both wild-type and mis mutants (G,H). By 8 h.p.f., endodermal cells extending into the hypoblast layer express cas in a “salt and pepper” distribution in wild-type (I, bracket). However, cas expression in mis mutant embryos at this stage is restricted to the blastoderm margin (J, arrowhead). (K,L) Closer analysis of cas gene expression in the YSL and DEL was carried out by FISH with a cas antisense probe (red) and DAPI labeling (blue), followed by confocal microscopy. The YSL layer can be distinguished from the DEL by the less dense arrangement of its nuclei and their larger size, and a white line has been drawn between the two layers. In wild-type embryos at this stage (K), expression of cas in the YSL is subsiding (asterisks indicate YSL nuclei associated with cas mRNA), while strong expression has begun in interspersed DEL cells tightly adjoining the YSL (arrows), the presumed endodermal precursors. On the other hand, mis embryos (L) show abnormally strong and persistent cas expression in the YSL and very few, if any, cas-expressing cells in the DEL. Stages are 5 h.p.f. (A–D, G,H), 6 h.p.f. (E,F), and 8 h.p.f. (I–L). (A,B,G,H) are lateral views with dorsal to the right, when identifiable. (C–F) are animal pole views (expression in embryos corresponding to these panels is largely symmetric and slight apparent asymmetries in the embryos shown are caused by photographing artifacts). (I,J) are dorsal views. (K,L) are optical sections through the embryonic blastoderm at about 1/3 of the way between the margin and the animal pole. EXPRESSION / LABELING:
|

Nodal-dependent activation of sox17 requires maternally-derived mis function. Wild-type (A,C,E,G,I,K) and mis mutant (B,D,F,H,J,L) embryos were injected at the 1-cell stage with cyc (A–D,G–J), sqt (E,F) or cas (K,L) mRNA, and assayed by in situ hybridization for the expression of no tail (A–D), gsc (E,F), cas (G,H) and sox17 (I–L). (A,B) In both wild-type and mis mutant embryos injected with cyc mRNA, ntl is ectopically induced in the blastoderm at 5 h.p.f. By 7 h.p.f., however, ntl ceases to be expressed in injected wild-type embryos (C), but remains strongly expressed in mis mutants (D). (E–H) Activation of nodal signaling results in ectopic gsc (E,F) and cas (G,H) expression at similar levels in both wild-type and mis mutant embryos, but leads to the strong induction of sox17 only in wild-type embryos (I) and not in mis mutants (J). (K,L) Overexpression of cas results in sox17 expression in wild-type (K) but not mis mutant (L) embryos. All panels are side views, except (E,F), which are animal views. |

Nodal-dependent induction of endoderm-specific genes in blastoderm explants is dependent on maternal mis function. (A) Experimental manipulation: cyc mRNA was injected at the 1-cell stage, and the blastoderm was manually excised away from marginal cells and the yolk cell at the late blastula stage (sphere stage, 4 h.p.f.), as originally carried out by David and Rosa (2001). At 7 h.p.f., explants from wild-type embryos show robust sox17 expression (B), while explants from mis mutant embryos lack sox17 expression (C). Embryos shown are a subset of representative embryos from two experiments with at least 15 wild-type and 15 mis embryos per experiment. Embryonic explants were only included if they were intact after dissection. Embryo diagrams in (A) are adapted from Kimmel et al. (1995) (images courtesy of ZFIN). |
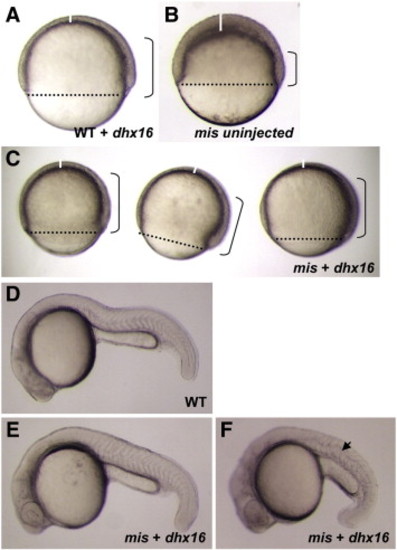
Rescue of the maternally mutant mis phenotype with wild-type dhx16 mRNA. (A–C) Live embryos observed at 8 h.p.f., corresponding to 75% epiboly stage in unaffected wild-type embryos. Dotted line shows the blastoderm margin, indicative of the front of epiboly; a vertical solid white line indicates the thickness of the animal-most cell layer, which normally thins out during gastrulation; and brackets demarcate the approximate extent of the involuted cell layer. (A) Wild-type embryos injected with dhx16 mRNA appear normal, shown here during gastrulation. (B) Control uninjected mis mutant embryos show morphogenetic defects characteristic of the mutation, including a reduction in epiboly, a thickened animal cap and reduced involution. (C) Three mis mutant embryos injected with dhx16 mRNA at the 1-cell stage, showing largely normal morphogenesis during gastrulation. (D–F) Live embryos observed at 24 h.p.f. (D) Wild-type embryo at 24 h.p.f. (E–F) mis mutant embryos with injected dhx16 mRNA survive gastrulation. A majority of these embryos exhibit a wild-type morphology (E), indicative of full genetic rescue. Some embryos, likely those with partial genetic rescue, survive gastrulation but have a shortened axis (F). Focal plane in (F) allows observing an abnormal undulation of the notochord (arrowhead), which may reflect remaining defects in dorsal convergence. As expected, uninjected mis mutant sibling embryos lyse during epiboly (not shown). Side views, animal pole to the top and dorsal side to the right in (A–C), and anterior to the left and dorsal to the top in (D–F). |
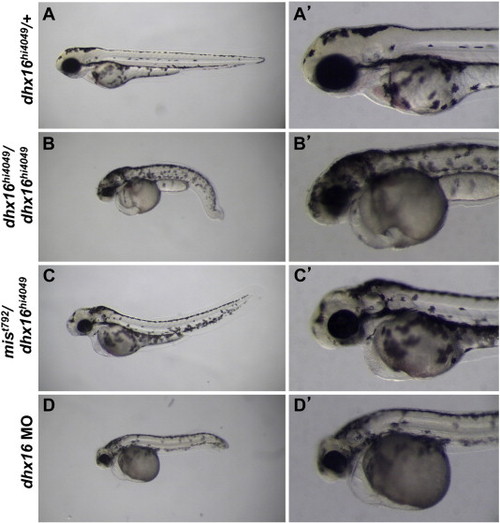
Complementation analysis and functional reduction of mis/dhx16. Live embryos at day 3 p.f. (A,A′) heterozygous for the dhx16hi4049 mutation have a wild-type morphology. (B,B′) Homozygous dhx16hi4049/dhx16hi4049 embryos exhibit necrosis and overall reduction in the head region, as well as a shortened and bent body axis. (C,C′) Necrosis, head reduction and body shape defects are present in mist792/dhxhi4049 transheterozygotes, albeit not to the extent found in dhx16hi4049 homozygotes. (D,D′) Embryos injected with 0.25–1 ng (shown here with 0.25 ng) of morpholino-conjugated oligonucleotide directed against the dhx16 translational start site develop a phenotype similar to that of dhx16hi4049/dhx16hi4049 homozygotes. Embryos injected with similar amounts of a control morpholino appear normal (not shown). In (B–D), the necrosis phenotype could be observed as early as day 2 p.f. as a darkening in the head (not shown), developed at day 3 p.f. into the smaller head (sometimes still associated with darkening) shown in these panels, and the embryos invariably lyse between days 3 and 6 of development. Side views, anterior left, dorsal up. Right column is a higher-magnification view of the left column focusing on the head region. PHENOTYPE:
|

The nuclear and cell division cycle, as well as the f-actin enrichment at the YSL, appear normal in mis mutant embryos. Wild-type (A,C,E) and mis mutant (B,D,F) embryos were fixed during epiboly (65% epiboly (A,B), 75% epiboly (C–F)) and labeled with phalloidin (green) and DAPI (blue) to detect f-actin and DNA, respectively. No obvious defects are observed with regards to the nuclear and cell division cycles, as well as in the formation of cell adhesive membranes and the f-actin network within the YSL (arrows). (A,B) are animal views and (C–F) are side views centered on the blastoderm margin. (E,F) are higher magnifications of the panels shown in (C,D), respectively, with focus on the membranes of the EVL. |

Gene expression markers of the hypoblast layer show reduced morphogenetic movements in maternal mis mutant (B,D,F) compared to wild-type (A,C,E). Embryos fixed at 85% epiboly (8.5 h.p.f.) and labeled to detect lefty2 (A,B), cyclops (C,D), and no tail (E,F) mRNA by in situ hybridization. In mutant embryos, the extension of the dorsal expression for these genes (double-headed arrows in A–E, a dorsal domain is not observed in F) is reduced along the anteroposterior axis, consistent with reduced involution of the hypoblast. Where not obscured by marginal expression, the dorsal domain is wider in mutants (brackets in A–D), consistent with defects in dorsal convergence. |

The microtubule network associated with the yolk cell during epiboly appears normal in maternally mutant mis embryos. Wild-type (A,A′) and maternal mis mutant embryos fixed at 30% epiboly (4.7 h.p.f.) and labeled to detect microtubules (green in A,B; grayscale in A′,B′) and DNA (blue in A,B). Arrays of microtubules extending towards the yolk cell can be observed in both types of embryos. Lateral views focusing on the blastoderm margin. |
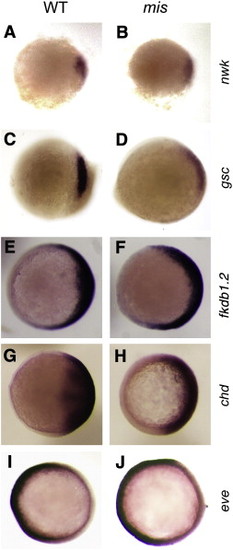
Spatial distribution of dorsal- and ventral-specific genes appears normal in maternally mutant mis embryos. Wild-type (A,C,E,G,I) and mis mutant (B,D,F,H,J) embryos exhibit normally restricted dorsal expression of nieuwkoid (A,B), goosecoid (C,D), foxb1.2 (E,F), and chordin (G,H), as well as normally restricted ventral expression of even skipped (I,J). (A–D) are embryos at 5 h.p.f., (E–F) at 7.5 h.p.f., and (G-J) at 8 h.p.f. All panels are animal pole views with dorsal to the right. Even though goosecoid expression is normally restricted, its levels of expression often appear reduced compared to wild-type. We have not further explored this effect, although it may be related to a requirement of mis function in the activation of additional nodal target genes such as goosecoid (Chen and Schier, 2001). |

Maternal mis function is not required for the activation of target genes by ectopic Wnt/Β-catenin and FGF signaling. Embryos were assayed for goosecoid expression at 70% epiboly (8 h.p.f.; A–F) or chordin expression at 30% epiboly (5 h.p.f.; G–J). Uninjected wild-type (A,G) and mis (B,H) show dorsally restricted expression of gsc and chd. Wild-type embryos injected with ß-catenin (C,I) or fgf8a (E) mRNA show expanded domains of dorsal gene expression. mis mutant embryos injected with ß-catenin (D,J) or fgf8a (F) mRNA also show ectopic expression of dorsal genes, although, as noted in Supp. Fig. 4, the overall levels of gsc expression can be reduced. |

Expression of mis/dhx16 mRNA during the cleavage and early gastrulation stages. In situ hybridization to detect mis/dhx16 in wild-type (A–C) and maternally mutant mis embryos (D–F), at the indicated stages. mRNA is present soon after fertilization (A), indicative of its maternal origin, and similar levels of mRNA can be observed throughout the cleavage stages (B) and early gastrulation (C). Levels of mRNA do not appear affected in mutant embryos in any of these stages (D–F). |
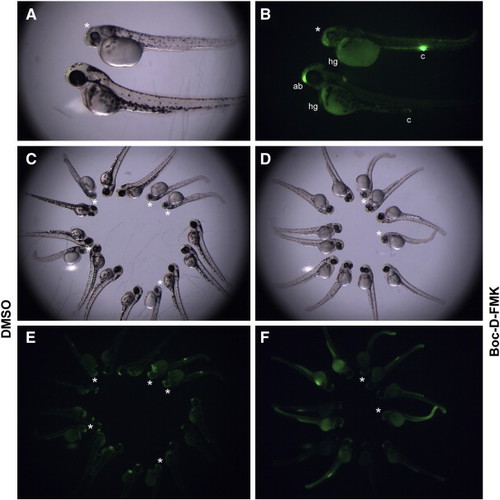
Loss of zygotic mis/dhx6 function in dhx16hi4049 homozygotes leads to caspase-dependent programmed cell death. (A,B) dhx16hi4049/dhx16hi4049 homozygote (top) and unaffected sibling (bottom) at day 3 p.f., shown under DIC optics (A) and fluorescence optics to detect acridine orange labeling (B). The darkening of the dhx16hi4049/dhx16hi4049 homozygotes (asterisk) correlates with an above-background number of acridine orange-labeled cells. (C–F) Groups of unsorted progeny from dhx16hi4049/+ heterozygotes, treated with either carrier solvent (C,E) or the general caspase inhibitor Boc-D-FMK (D,F) and visualized with DIC optics (C,D) and to detect acridine orange labeling (E,F). In control-treated clutches, the expected fraction (1/4) of embryos shows signs of necrosis associated with increased acridine orange labeling (asterisks). Exposure to caspase inhibitor reduces both the penetrance and expressivity of the necrosis phenotype, and these effects correlate with decreased acridine orange labeling. At this stage of development, several regions show acridine orange labeling (sensitive to caspase inhibitor) in control, wild-type embryos, such as the anteriormost region of the brain ((ab), see also Furutani-Seiki et al., 1996) and hatching gland (hg) cells overlying the yolk cell. Fluorescence can also be observed in the developing cloaca (c), although this labeling appears non-specific as it can be observed in the presence of caspase inhibitor (F). Acridine orange labeling in the anteriormost region of the brain, observed in wild-type embryos at this stage, appears reduced in mutant embryos (B), possibly because these embryos do not undergo the normal developmental events that lead to apoptosis in wild-type embryos in this region. |
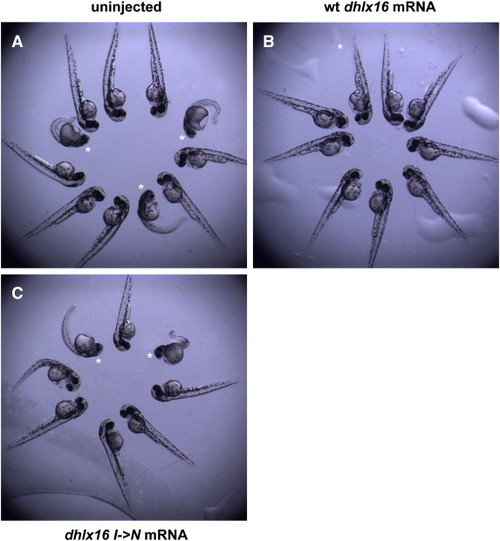
Expression of a wild-type, but not mutant, form of dhx16 rescues the necrosis associated with the dhx16 zygotic null phenotype. (A–C) Groups of unsorted progeny from two dhx16hi4049/+ heterozygous parents, uninjected (A), or injected with mRNA encoding the wild-type (B) and maternal-effect mutant (mist792) (C) alleles of dhx16. The necrosis phenotype observed in uninjected controls, expected in ¼ of embryos (experimentally observed in 26.0% of embryos, n = 100; asterisks in (A)) is not observed (0%, n = 29) in embryos expressing wild-type Dhx16 (B). Embryos injected with mRNA coding for the product corresponding to the maternal-effect mist792 allele (Dlhx16 I->N product) exhibit necrosis embryos at a fraction (25.5%, n = 47) and with a similar phenotypic strength as uninjected siblings (asterisks in (A,C)). Data are compiled from two separate experiments with similar results. |
Reprinted from Developmental Biology, 353(2), Putiri, E., and Pelegri, F., The zebrafish maternal-effect gene mission impossible encodes the DEAH-box helicase Dhx16 and is essential for the expression of downstream endodermal genes, 275-289, Copyright (2011) with permission from Elsevier. Full text @ Dev. Biol.