
- Title
-
ELABELA: A Hormone Essential for Heart Development Signals via the Apelin Receptor
- Authors
- Chng, S.C., Ho, L., Tian, J., and Reversade, B.
- Source
- Full text @ Dev. Cell
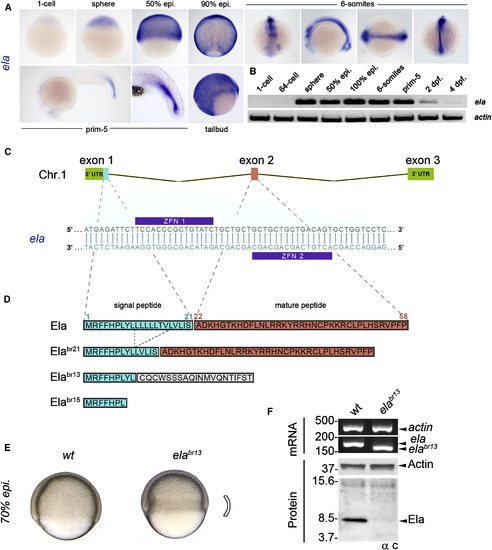
Generation of an Allelic Series of Mutant ela Zebrafish (A) By WISH in zebrafish embryos, ela is found to be zygotically expressed and ubiquitous in the blastoderm. During gastrulation, its expression becomes axial and is strongest in the neural tube. (B) RT-PCR analysis shows that, relative to actin, expression of ela is exclusively zygotic, peaking at 100% epiboly and becoming absent by 4 dpf. (C) In zebrafish, ela consists of three exons located on chromosome 1. A custom pair of ZFNs targeting exon 1 of ela was used to create an allelic series of mutations within the signal peptide of Ela. (D) Three distinct loss-of-function alleles were generated. The elabr21allele results in a unique 7 aa in-frame deletion in Ela’s signal peptide. Alleles elabr13 and elabr15 caused premature stop codons and disrupted the reading frame, resulting in no Ela mature peptide. (E) Relative to WT embryos, homozygous null elabr13 embryos show defective epiboly movements and a constricted germ ring at the involuting margin at 70% epiboly. (F) RT-PCR reveals that at 100% epiboly, endogenous ela and elabr13 mRNAs are of a distinct size. Western blotting using the α C antibody shows that endogenous Ela is recognized in WT embryos and is absent in null elabr13 siblings. |

ela Knockout Zebrafish Have Severe Cardiovascular and Endoderm Defects (A) Null elabr21 larvae display pericardial edema (white arrowheads) and accumulated erythrocytes (red arrowheads), and have no blood circulation. Variable posterior anomalies are observed, including loss of ventral fin (black arrowheads), tailbud duplications (inset), and extreme tail/trunk truncations (bottom embryo). (B) Loss of ela causes severe cardiac anomalies ranging from mild heart dysplasia to total heart agenesis, as shown by hematoxylin and eosin staining on sections from the top and bottom embryos in (A). (C) Null elabr13 fish have a severe reduction of cmlc1 expression, which marks the developing heart. (D) Null elabr13 fish display increased hematopoiesis as judged by the upregulation of scl, a marker of blood precursors. (E) Classification of null elabr13 larvae with varying degrees of tail defects. Class I larvae are defined as having pericardial edema and tail blood clot. Class 2 larvae have ventral fin defects or tailbud duplications in addition to class 1 phenotypes. Class 3 larvae have all phenotypes of class 1 combined with mild to severe tail/trunk truncations. (F) Percentages of ela mutant fertile adults that were obtained from heterozygous intercrosses using all three alleles. (G) Relative to WT embryos, homozygous null elabr13 embryos show convergence-extension defects resulting in delayed blastopore closure and thickened notochord (insets) as indicated by altered bra expression. (H) The loss of ela causes defective migration of mediolateral gata5-expressing cells, which mark mesendoderm cells that guide heart progenitors to the anterior lateral plate mesoderm. (I) ela mutant embryos show reduced foxa2 expression compared with WT embryos. (J) ela mutant embryos show a more compact sox17 expression pattern compared with WT embryos, whereas sox17+ forerunners cells are not affected. (K) ela null embryos have approximately 40% less sox17+ cells than their heterozygous siblings at 75% epiboly. (L) At 75% epiboly, elabr21 and elabr15 mutant embryos exhibit sox17 expression defects similar to those observed for elabr13 (see also Figure S1). EXPRESSION / LABELING:
PHENOTYPE:
|

Ela’s Cognate Receptor for Endoderm Differentiation Is Aplnr (A) ELA and apelin are very basic hormones with isoelectric points exceeding 12. (B) By qPCR, the onset of transcription of aplnra and aplnrb coincides with that of ela at the MBT; apln expression debuts 5 hr later during gastrulation (data are represented as mean ± SEM). (C) Relative to control embryos, the expression of aplnra and aplnrb at 70% epiboly becomes stronger and confined to the most equatorial hypoblast in the ela mutant (white arrowheads: specific aplnra expression in the animal pole). (D) WT embryo with a beating heart and blood circulation at 6 dpf, normal sox17 expression in definitive endoderm at 75% epiboly, no erythrocyte accumulation in the ICM, and cmlc1 expression in the heart-forming region at 30 hpf. (E) aplnr morphants phenocopy ela mutant embryos, with no beating heart, loss of blood circulation at 6 dpf, reduced sox17 expression in definitive endoderm at 75% epiboly, accumulation of erythrocytes in the ICM, and loss of cmlc1 expression in the heart-forming region at 30 hpf. (F) ela mutant embryos are nearly indistinguishable from Aplnr morphants and have no beating heart or blood circulation at 6 dpf, show reduced sox17 expression in definitive endoderm at 75% epiboly, have accumulated erythrocytes in the ICM, and no cmlc1 expression in the heart-forming region at 30 hpf. (G) In 293T cells, overexpression of zebrafish aplnra or aplnrb, or human APLNR is sufficient to confer cell-surface binding to recombinant AP-ELA. (H) In 293T cells, overexpression of zebrafish aplnrb, but not its mutant form (grinch) carrying the W90L missense mutation, or GPR15 (an orphan GPCR closely related to APLNR), is enough to afford binding to recombinant AP-ELA. EXPRESSION / LABELING:
PHENOTYPE:
|
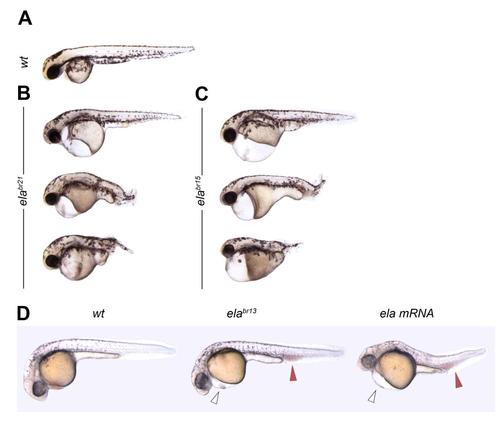
Homozygous elabr21 and elabr15 Mutants and ela overexpressing Fish Have Similar Phenotypes, related to Figure 3 (A) Wildtype larvae at 6 days post-fertilization. (B) The 7 amino-acid in frame mutant elabr21 allele causes identical phenotypes as the two frameshift mutants elabr13 and elabr15. (C) The elabr15 mutant fish are indistinguishable from the elabr21 and elabr15 alleles. (D) Overexpression of 200 pg of zebrafish ela ORF mRNA causes heart dysgenesis (open arrows) and accumulation of erythrocytes in the ICM (red arrows), similar to ela null fish. PHENOTYPE:
|
Reprinted from Developmental Cell, 27(6), Chng, S.C., Ho, L., Tian, J., and Reversade, B., ELABELA: A Hormone Essential for Heart Development Signals via the Apelin Receptor, 672-680, Copyright (2013) with permission from Elsevier. Full text @ Dev. Cell