
- Title
-
Yap-lin28a axis targets let7-Wnt pathway to restore progenitors for initiating regeneration
- Authors
- Ye, Z., Su, Z., Xie, S., Liu, Y., Wang, Y., Xu, X., Zheng, Y., Zhao, M., Jiang, L.
- Source
- Full text @ Elife
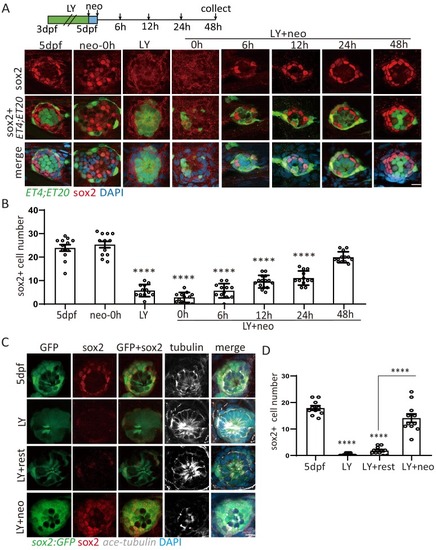
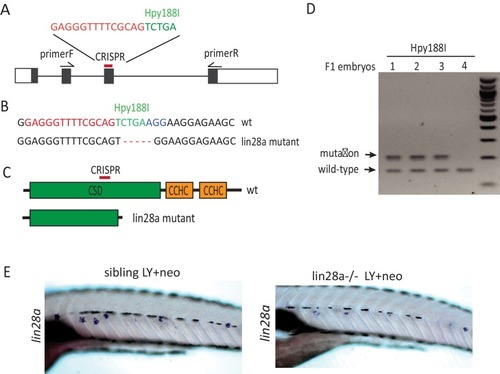
(A, B) ET4;ET20 larvae were treated with neomycin, LY411575 (3dpf-5dpf), or neomycin following LY (LY+neo), and collected at indicated time points post neomycin treatment for sox2 immunostaining. The number of sox2+ progenitors was not affected post neo, while it was significantly decreased in LY and LY+neo-0h. The sox2+ progenitors were regenerated post LY+neo and recovered to normal level at 48 hr post LY+neo. (C, D) The sox2:GFP reporter was treated with LY from 3dpf to 5dpf to exhaust GFP+ progenitors. GFP+ progenitors cannot be regenerated when resting in normal medium for 2 days post LY treatment (LY+rest). In contrast, sox2+ progenitors were quickly recovered to normal level at 2-day post LY+neo. Scale bar equals 10 μm. All groups are compared with 5dpf unless indicated |

|
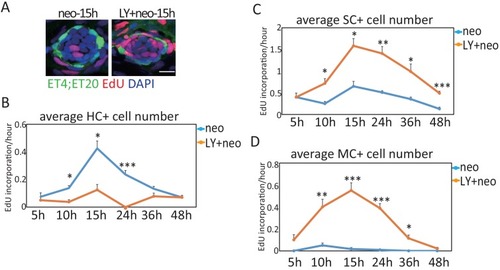
Larvae treated with neo or LY+neo were incorporated with EdU for different time points and counted for numbers of differentiating cells (EdU+ |

( |
( |
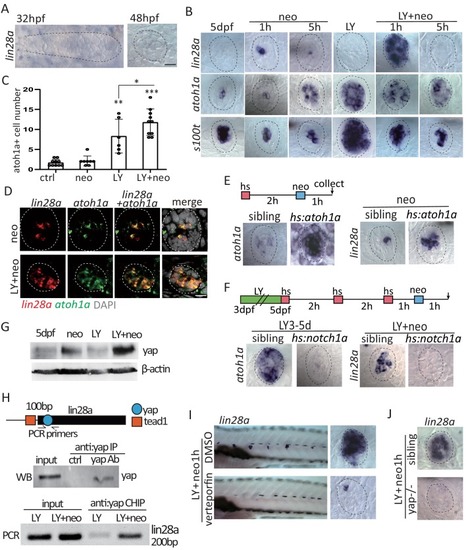
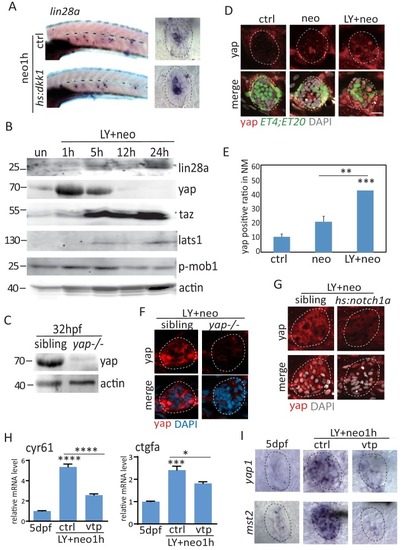
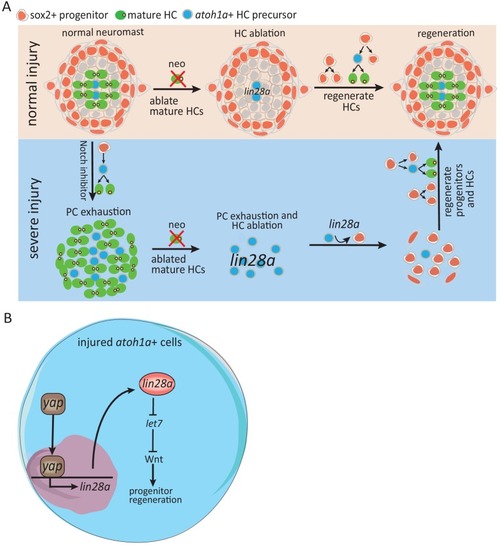
(A) Lin28a was not expressed in the developing lateral line primordium or neuromast. (B) Treatment with Notch inhibitor LY411575 from 3dpf to 5dpf increased expression atoh1a and s100t. More lin28a expression was observed in LY+neo-1h compared with neo-1h. No lin28a was detected at 5 hr post LY+neo or neo. (C) The number of atoh1a-transcribed cells detected by in situ were higher at 1 hr post LY+neo compared with LY. (D) Double fluorescent in situ showed that lin28a was co-expressed with atoh1a post neo or LY+neo. (E) Induction of lin28a post injury was increased when atoh1a was overexpressed in hs:atoh1a. (F) Lin28a expression was completely blocked when atoh1a was inhibited in hs:notch1a. (G) Western blot results showed that LY+neo induced higher yap expression compared with neo alone. (H) Motifs of yap and tead1 (co-transcriptional factor of yap) binding sites were predicted near lin28a transcriptional start site. CHIP-PCR results verified that yap directly binds the predicted motif. (I and J) Inhibition of yap using verteporfin or yap mutant blocked lin28a induction post LY+neo. Scale bar equals 10 μm. EXPRESSION / LABELING:
PHENOTYPE:
|
( |
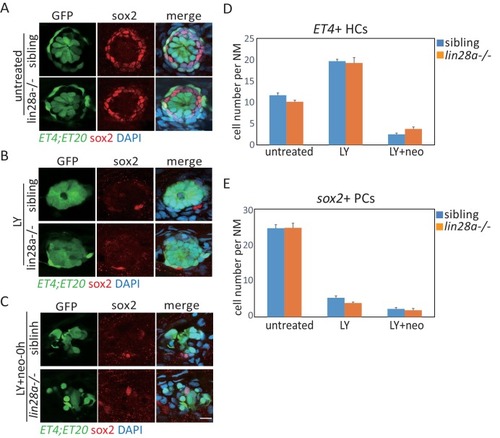
We compared numbers of |
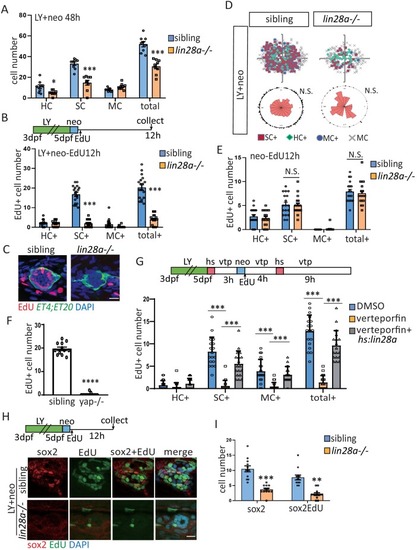
(A) The number of SCs and total cells were counted at 48 hr post LY+neo and were significantly decreased in lin28a mutant compared with sibling. (B–E) The ET4;ET20 larvae were incorporated with EdU post neo or LY+neo treatment. Three populations labeled with ET4+EdU+ (HC+), ET20+EdU+ (MC+) or ET4-ET20-EdU+ (SC+) were counted and recorded with location information. The proliferative SCs post LY+neo were significantly decreased in lin28a mutant compared with sibling (B and C), while not changed in neo-induced regeneration (E). (D) EdU plots show the positions of EdU+ nuclei of 18 neuromasts superimposed on the same plane, and rose diagrams document the angular positions of SC+. The results show that the proliferative SCs are evenly distributed in each quadrant with no polarization post LY+neo. (F) LY+neo-induced Edu incorporation was significantly reduced in yap mutant. (G) The hs:lin28a larvae were heat-shocked and pre-treated with 10 μM verteporfin before adding neomycin. Samples were collected at 14 hr post LY+neo for EdU+ cell counting. Proliferation (SC+) is significantly decreased post LY+neo in verteporfin, which could be rescued by overexpression of lin28a with hs:lin28a. (H, I) The numbers of regenerated sox2+ progenitors (sox2) and proliferative progenitors (sox2EdU) post LY+neo were both reced in lin28a mutant. Scale bar equals 10 μm. PHENOTYPE:
|

(A–D) ET4;ET20;hs:lin28a larvae were incorporated with BrdU for 24 hr following heat-shock and/or neomycin treatment. (A, B) Overexpression of lin28a increased number of proliferative SCs (SC+) in both untreated and neomycin conditions. (C, D) BrdU plots and rose diagrams indicate that locations of SC+ still remain dorsally and ventrally polarized in hs:lin28a. (E, F) OverexpressUion of lin28a is sufficient to partially restore the exhausted sox2+ progenitors post LY. Scale bar equals 10 μm. PHENOTYPE:
|

The |

(A–B) Larvae were stained with sox2 antibody and atoh1a RNA probe at 6 hr post neo or LY+neo treatment. The number of atoh1a+sox2+ cells was significantly increased in LY+neo group. (C–D) The atoh1a:TdTomato larvae were used to trace atoh1a+ HC precursors in ctrl, LY or LY+neo. Results showed that Tomato+ cells labeled partial ET4+ HCs and ET4-sox2- HC precursors (yellow arrows). However, many atoh1a+ cells start to express sox2 from 12 hr post LY+neo and their numbers were significantly increased compared with normal larvae. The arrowheads in (C) pointed the atoh1a+sox2+ cells at 48 hr post LY+neo. (E) The atoh1a:TdTomato;hs:lin28a larvae was treated with LY+neo and verteporfin and collected for immunostaining with sox2 antibody. The cell number of Tomato+sox2+ is decreased in verteporfin and overexpression of lin28a could rescue the phenotype. (F–G) The LY-treated hs:lin28a larvae were heat-shocked to overexpress lin28a. Samples were collected for staining with sox2 antibody and atoh1a RNA probe. The number of atoh1a+sox2+ cells was significantly increased in hs:lin28a group, indicating that lin28a is sufficient to express sox2 in atoh1a+ HC precursors. Scale bar equals 10 μm. |

( |
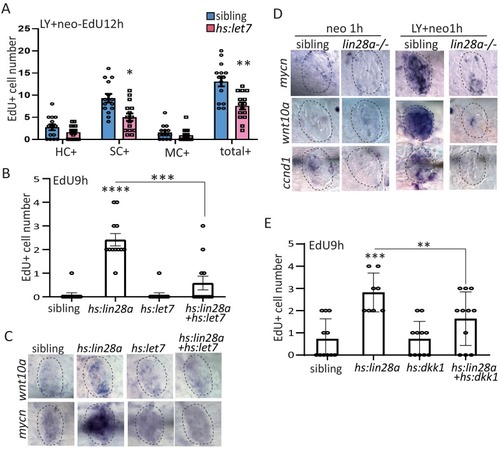
(A) We created hs:let7 transgenic line and found that it recapitulates the phenotype of lin28a-/- by decreasing proliferative SCs post LY+neo. (B) The induction of EdU+ proliferative cells was blocked by hs:let7, indicating that let7 microRNA acts downstream of lin28a to induce proliferation. (C) In situ hybridization results showed that expression of Wnt pathway genes, such as wnt10a and mycn, were increased in hs:lin28a. The activation of Wnt pathway genes in hs:lin28a were blocked when let7 was overexpressed, indicating that let7 acts downstream of lin28a to inhibit Wnt pathway. (D) Expressions of Wnt pathway genes, such as mycn, wnt10a and ccnd1 (cyclind1), were highly induced at 1 hr post LY+neo in sibling, but were not detected in lin28a-/-. (E) Inhibition of Wnt pathway with hs:dkk1 decreased lin28a-induced EdU+ proliferative cells, indicating that Wnt activation acts downstream of lin28a to induce regeneration. |

In situ results show that expression of |
( |