
- Title
-
Distinct Wnt signaling pathways have opposing roles in appendage regeneration
- Authors
- Stoick-Cooper, C.L., Weidinger, G., Riehle, K.J., Hubbert, C., Major, M.B., Fausto N., and Moon, R.T.
- Source
- Full text @ Development
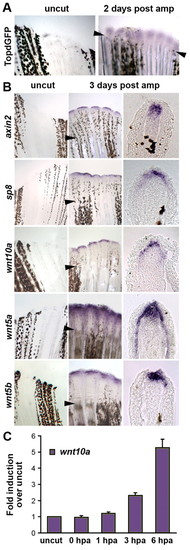
Wnt/β-catenin signaling is upregulated in regenerating zebrafish tail fins. (A) Wnt/β-catenin reporter (TOPdGFP) activity, detected by in situ hybridization for GFP RNA (blue), is upregulated in the blastema of regenerating fins of zebrafish homozygous for the transgene at 48 hpa (n=5; arrowheads indicate the amputation plane). Control is a non-amputated TOPdGFP fin. At 3 dpa (n=3) and 5 dpa (n=3), TOPdGFP was still upregulated (not shown). (B) In situ hybridization of control non-amputated fins (left panels), regenerating fins at 3 dpa (middle panels), and cross-sections of fins at the same stage (right panels). The Wnt/β-catenin target genes axin2 and sp8 are expressed in the distal tip of the blastemal mesenchyme and in the basal epithelial layer of the regeneration epidermis, respectively. wnt10a is expressed in the distal tip of the blastema. Both wnt5a (for nomenclature, see Fig. S2 in the supplementary material) and wnt5b are expressed in the basal epithelial layer of the regeneration epidermis and in the distal tip of the blastema, with wnt5a extending far proximally in the basal epithelium. (C) wnt10a expression levels in uncut control and regenerating fins at 0 hpa (sample isolated immediately after fin amputation), 1 hpa, 3 hpa and 6 hpa as determined by quantitative PCR. RNA was isolated from the tips of fins of 10 wild-type fish for each time point. Expression levels were normalized to β-actin levels (normalization to 18S rRNA levels produced very similar results) and fold-induction calculated by setting the level of uncut fins to 1. Quantitative PCR was performed four times on the same samples; error bars represent the s.e.m. EXPRESSION / LABELING:
|

Wnt/β-catenin signaling is required for zebrafish tail fin regeneration. (A) Experimental scheme. Tail fins were amputated from wild-type, hsDkk1GFP or hsΔTcfGFP transgenic zebrafish and heat shocks were applied twice daily for the time periods indicated by the colored lines. (B) Continuous suppression of Wnt target gene expression in hsΔTcfGFP (n=15), or reduction of Wnt/β-catenin signaling in hsDkk1GFP transgenic fish (n=19), for 7 days starting shortly before amputation inhibits fin regeneration. Live fins were photographed at 1 dpa (left panels) and 7 dpa (right panels). (C) Overexpression of Dkk1 starting at 1 dpa inhibits fin regeneration (left panel; n=18); overexpression from 3 dpa results in partial inhibition of regeneration (right panel; 11 of 15 fins). Live fins were photographed at 7 dpa; corresponding wild-type controls regenerated normally (not shown). PHENOTYPE:
|
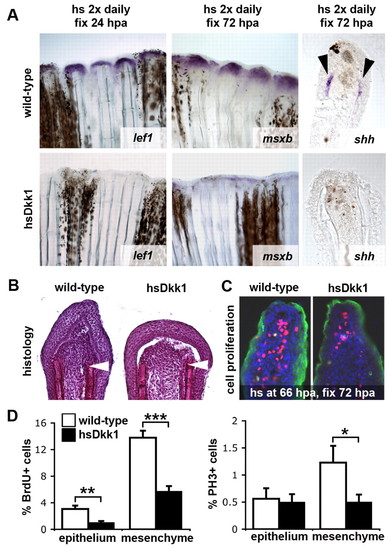
Wnt/β-catenin signaling regulates specification and proliferation of the regeneration blastema. (A) Expression of lef1, a marker for the basal epidermal layer of the regeneration epithelium, msxb, marking the mesenchymal progenitor cells of the blastema, and shh, expressed in basal epidermal cells (shown in thick sections), is strongly reduced in Dkk1-overexpressing fins. lef1 is shown at 24 hpa (n=4), msxb (n=4) and shh (n=4) at 72 hpa. Fish were heat shocked twice daily starting shortly before amputation. (B) Hematoxylin-stained sections of tail fin regenerates at 48 hpa. Dkk1-overexpressing fins (right panel; n=6) display reduced numbers of deep mesenchymal cells of the blastema. Fish were heat shocked twice daily starting shortly before amputation. Arrowheads indicate the plane of amputation. (C) 72 hpa regenerates stained for BrdU (red), phosphorylated histone H3 (PH3, green) and DAPI (blue). Cell proliferation in both the mesenchyme and epithelium is decreased in Dkk1-overexpressing fins. Fish were heat shocked once at 66 hpa and fixed at 72 hpa. (D) Quantification of the cell proliferation defects in Dkk1-overexpressing regenerating fins. The fraction of BrdU-positive (left) and PH3-positive (right) cells relative to the total number of cells (DAPI-positive) is shown in percent (n=11). Error bars represent the s.e.m; *P=0.0495; **P=0.0025; ***P=7.076x10-6 (two-tailed). |

β-catenin-dependent and β-catenin-independent Wnt signaling pathways have opposing roles in zebrafish fin regeneration. (A,D) Overexpression of Wnt8 in hsWnt8GFP transgenic fish induces the Wnt/β-catenin target gene axin2 in regenerating fins 6 hours after heat shock at 3 dpa (A; 3 of 4 fins), whereas overexpression of Wnt5b in hsWnt5bGFP transgenic fish represses axin2 expression (D; n=4). Note that staining reactions were stopped as soon as a robust signal could be detected in most samples of one experimental group. Robust signal was first detected in hsWnt8GFP fins (versus wild-type controls) after a short amount of time and reactions were stopped (A), whereas robust signal was first detected in wild-type controls (versus hsWnt5GFP fins) after a longer staining reaction (D), thus accounting for the difference in wild-type signal between groups. (B,E) Cell proliferation in regenerating fins, as detected by BrdU incorporation and staining with an anti-PH3 antibody, is increased by overexpression of Wnt8 (B; n=14), and repressed by overexpression of Wnt5b (E; n=10). Fish were heat shocked once at 66 hpa and fixed at 72 hpa. The percentage of BrdU-positive or PH3-positive cells relative to the total number of cells in sections of regenerating fins is shown. Error bars represent the s.e.m. (C,F) Whereas overexpression of Wnt8 for 10 days starting shortly before amputation has no obvious effect on overall length of the regenerate (C; n=16), overexpression of Wnt5b completely inhibits regeneration (F; n=16). (B) *P=0.0579; **P=0.0082; ***P=0.0002 (two-tailed). (E) *P=0.0377; **P=0.0123; ***P=0.0006 (two-tailed). |
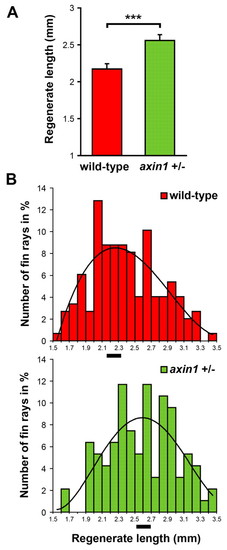
Fins regenerate faster in fish heterozygous for a loss-of-function mutation in axin1. (A) Average length of regenerating tail fins at 7 dpa is increased in fish heterozygous for an axin1 loss-of-function mutation (mbltm013) compared with wild-type siblings. Results of one representative experiment of three are shown. To determine the length of the regenerate for individual fish, the average length of the third, fourth and fifth dorsal regenerating fin ray was calculated. n=12 wild-type fish and 9 mbl heterozygous fish; error bars represent the s.e.m. of the average regenerate lengths; ***, P=0.0009 (one-tailed). (B) The number of fin rays (in percent of the total number counted) is plotted against the length of the regenerate (in 0.1 mm intervals) for wild-type (upper bar graph) and axin1 heterozygous fish (lower bar graph). The curves represent a fifth-order polynomial trendline. The average regenerate length is marked by black bars at the x-axis. 148 fin rays were counted (combined results from three experiments) in 19 wild-type fish and 94 rays were counted in 10 axin1+/- fish. PHENOTYPE:
|
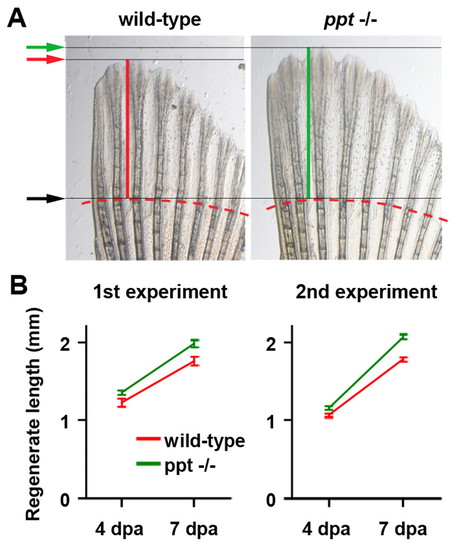
Fins regenerate faster in wnt5b mutant fish. (A) Dorsal half of regenerating tail fins of wild-type and wnt5b (ppt) homozygous mutant fish at 7 dpa. The amputation plane is indicated by a dashed red line, the length of the third fin ray by red or green bars and arrows. Note that the regenerate is longer in ppt than in wild-type fish. (B) The average length of the regenerate of wild-type and ppt mutant fish at 4 and 7 dpa in two independent experiments. To determine the length of the regenerate for individual fish, the average length of the third, fourth and fifth dorsal regenerating fin ray was calculated. Experiment 1: n=14 wild-type, 12 ppt fish. Experiment 2: n=12 wild-type, 11 ppt fish. Error bars represent the s.e.m. of the average regenerate lengths. For P values, see Table S2 in the supplementary material. Note that absolute fin lengths cannot be compared between experiments because water temperatures and thus regenerative speed and exact times of photography varied between experiments. PHENOTYPE:
|
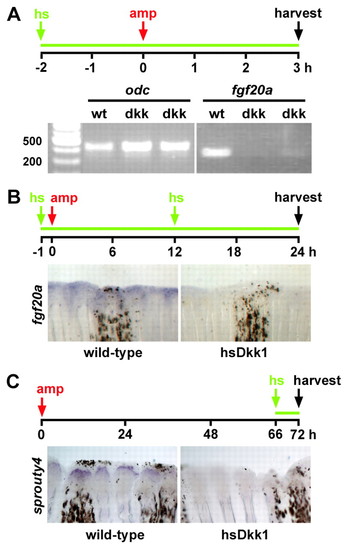
Wnt/β-catenin signaling regulates FGF signaling during fin regeneration. (A) fgf20a expression as detected by semi-quantitative RT-PCR is greatly reduced in Dkk1-overexpressing fins at 3 hpa. Wild-type and hsDkk1GFP transgenic fish were treated according to the schematic (hs, heat shock; amp, amputation; the green line indicates inhibition of Wnt/β-catenin signaling), and RNA was harvested from the tissue adjacent to the amputation plane of nine wild-type and two groups of nine hsDkk1GFP transgenic fins. odc1 amplification serves as a loading control. The experiment was repeated four times using two sets of biological samples and representative results are shown. (B) fgf20a expression is greatly reduced as detected by in situ hybridization in hsDkk1GFP transgenic fins (5 of 6 fins) at 24 hpa as compared with wild-type fins. (C) sprouty4 expression is greatly reduced 6 hours after heat shock in hsDkk1GFP fins (n=3) at 72 hpa. EXPRESSION / LABELING:
|
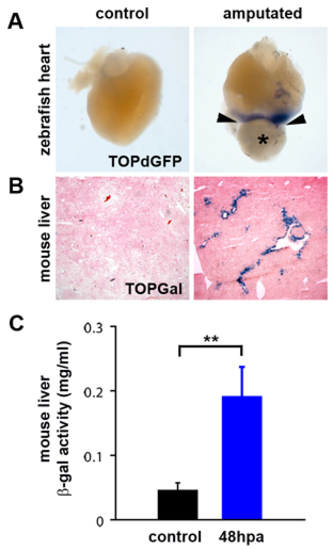
Wnt/β-catenin signaling is upregulated in regenerating zebrafish heart and mouse liver. (A) Wnt/β-catenin reporter (TOPdGFP) activity (detected by in situ hybridization for GFP RNA, blue) is upregulated in regenerating zebrafish hearts (n=4) adjacent to the amputation plane (arrowheads) at 3 dpa (*=blood clot). Control is a sham-operated TOPdGFP heart at 3 days post-sham. TOPdGFP is also expressed adjacent to the amputation plane at 7 dpa (n=3) and 14 dpa (n=3) (data not shown). (B) Wnt/β-catenin reporter (TOPGal) (DasGupta and Fuchs, 1999) activity (detected by staining for β-galactosidase activity, blue) is upregulated in periportal hepatocytes in regenerating mouse liver (n=8), 48 hours post-partial hepatectomy. (C) Quantification of TOPGal activity in regenerating mouse liver. β-galactosidase activity was measured in control lobes (resected lobes removed at the time of partial hepatectomy) and regenerating lobes, collected 48 hpa. Control lobes were compared to regenerating lobes harvested from the same animal. Asterisks indicate a statistically significant difference between experimental groups (n=8, P<0.01, Paired Student’s t-Test). Error bars represent the s.e.m. |

Establishment of heat-shock inducible Dickkopf1GFP and heat-shock inducible Wnt5bGFP transgenic zebrafish lines. (A) Maps of the transgenes used. (B) Strategy used to establish and characterize transgenic fish (see Materials and methods). (C,D) Heat-shock of heterozygous hsDkk1GFP transgenic embryos at 24 hpf for 2.5 hours results in ubiquitous expression of the Dkk1GFP fusion protein. (E,F) Heat-shock of heterozygous hsDKK1GFP transgenic adult fish for 1 hour causes expression of the fusion protein in amputated tail fins. (G-I) Heat-shock of heterozygous hsDkk1GFP transgenic embryos for 2 hours during gastrulation (starting at shield stage, 6 hpf) causes severe defects in anteroposterior patterning that closely resemble those caused by knockdown of both ORFs of wnt8 using translation-blocking morpholinos. Heterozygous carriers of the transgene were crossed to wild-type fish, and the resulting clutch of embryos heat shocked at 6 hpf for 2 hours. Wild-type and transgenic embryos were identified by GFP fluorescence, sorted and photographed at 24 hpf. (J,K) Overexpression of Dkk1GFP in embryos doubly transgenic for hsDKK1GFP and the reporter TOPdGFP is sufficient to suppress β-catenin-dependent expression of the TOPdGFP transgene. Heterozygous carriers of the hsDkk1GFP transgene were crossed to homozygous carriers of the TOPdGFP transgene. The resulting clutch of embryos was heat-shocked at 9 hpf for 1 hour, embryos fixed at 12 hpf and processed for two-color in situ hybridization with an EGFP probe detecting the TOPdGFP transgene in blue and a mmGFP probe detecting the dkk1GFP transgene in brown. The pictures show the head region of flat-mounted embryos, anterior left. Expression of TOPdGFP at this stage is primarily in the tectum and hindbrain. (L,M) Overexpression of Wnt5bGFP in embryos heterozygous for the hsWnt5bGFP transgene during gastrulation causes phenotypes typical for activation of β-catenin-independent Wnt signaling pathways, namely convergent-extension defects, resulting in short embryos with compressed somites and wavy notochord. Heterozygous carriers for the hsWnt5GFP transgene were crossed to wild-type fish, the resulting embryos heat shocked at 6 hpf for 2 hours, sorted into wild-type and transgenic carriers by GFP fluorescence and photographed at 24 hpf. PHENOTYPE:
|

wnt5b expression during fin regeneration is regulated by Wnt/β-catenin signaling. Fins of hsDkk1GFP transgenic and wild-type fish were amputated, fish heat shocked once at 72 hpa and fins fixed 6 hours later. wnt5b expression is downregulated in Dkk1-expressing fins (n=10). EXPRESSION / LABELING:
|