
- Title
-
The chemokine receptor CXCR4 promotes granuloma formation by sustaining a mycobacteria-induced angiogenesis programme
- Authors
- Torraca, V., Tulotta, C., Snaar-Jagalska, B.E., Meijer, A.H.
- Source
- Full text @ Sci. Rep.
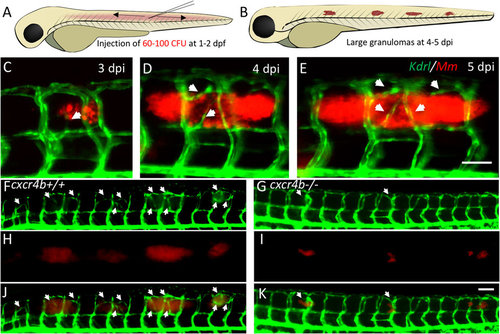
Trunk granuloma formation and induction of angiogenesis by M. marinum. (A,B) Schematic representation of injection location and of granuloma expansion in zebrafish embryos/larvae upon injection of Mm into the trunk. (C–E) Longitudinal imaging of vascular and bacterial growth during development of a trunk granuloma in the Tg(kdrl:eGFP) line at 3, 4 and 5 dpi. (F–K) Formation of trunk granulomas and their vascularisation in a cxcr4b+/+ and cxcr4b−/− larva at 5 dpi. White arrows indicate sites of abnormal vascularisation associated to bacterial growth, notably largely initiated in the wt but rare in the mutant. See also Fig. 2 for quantification. Scale bars: 100 μm. |
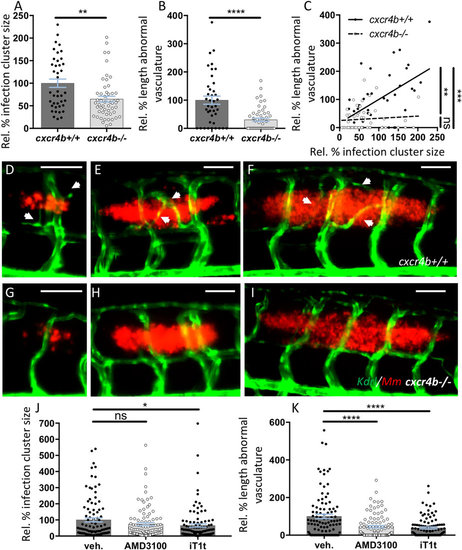
Cxcr4b mutation impairs granuloma-induced angiogenesis. (A,B) cxcr4b mutants display a reduced expansion rate of local granulomatous lesions (A), which coincided with the incapability to induce an angiogenic programme (B). (C) Vascularisation/granuloma expansion correlation analysis. In wt, the expansion of the granulomas depends on the activation of the angiogenic programme at the infection focus. Differently, in cxcr4b mutants, no significant correlation between granuloma size and vessel length could be observed and even large granulomas failed to initiate angiogenesis, suggesting that the differential activation of the angiogenic programme is not the effect, rather the cause, of reduced burden in mutants. Aberrant vasculature was measured as the two-dimensional length of vessels not found in uninfected larvae and that solely form in association to granulomas. Infection cluster size was measured as the two-dimensional area of individual trunk lesions. Values are expressed in percentage relative to average infected cxcr4b+/+ (set to 100%). Statistics in C indicates that the slopes of cxcr4b−/− and cxcr4b+/+ trend lines are significantly different (linear regression comparison test) and that in wt (but not in mutants) this slope is different from 0 (x axis, Pearson correlation test). Each data point in A-C refers to individual trunk granulomas at 5 dpi. Experiments were performed in 3 replicates (cumulated in the graphs), each with 4–12 larvae per group. Total number of granulomas analysed: 40 (cxcr4b+/+), 52 (cxcr4b−/−). (D–I) Representative images of comparably-sized 5 dpi trunk granulomas in cxcr4b−/− and cxcr4b+/+ (quantified in A–C). Notably cxcr4b−/− granulomas display a defect in local vascularisation, which is independent of the granuloma size. Scale bars: 100 μm. (J–K) Treatments with CXCR4 antagonists AMD3100 and IT1t phenocopy cxcr4b mutants and display attenuated (IT1t) expansion of granulomatous lesions (J) and reduced (AMD3100 and IT1t) induction of granuloma-associated angiogenesis (K). Experiments in J–K were performed in Tg(fli1a:eGFP) background. Data are analysed as in Fig. 2A,B. Experiments were performed in 2 replicates (cumulated in the graphs). Total number of granulomas analysed: 101 (vehicle), 101 (AMD3100), 103 (IT1t). |
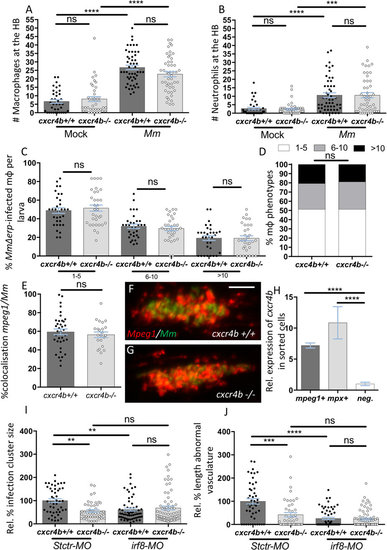
cxcr4b is not required for the establishment of macrophage parasitism, but macrophages are necessary to mediate cxcr4b-dependent angiogenesis. (A,B) Recruitment of macrophages (A) and neutrophils (B) 3 hours post local hindbrain (HB) infection at 2 dpf with Mm was unaffected by cxcr4b mutation. Each data point in (A,B) represents the recruitment measured in 1 embryo. Experiments were performed in 2 replicates (cumulated in the graphs), each with 17–35 embryos per group. Total number of embryos analysed: 38 (Mock-cxcr4b+/+), 41 (Mock-cxcr4b−/−), 53 (Mm-cxcr4b+/+), 49 (Mm-cxcr4b−/−). C–D cxcr4b is dispensable for the capability of macrophages to counteract intracellular replication of Mm Δerp. Infected macrophages (mpeg1:mCherry-F+) were classified into three phenotypic classes, according to the severity of intracellular infection (1–5, 6–10, or >10 bacteria). (C) represents the percentage of macrophages per larva that populates each class. (D) represents the overall distribution of macrophage phenotypes (macrophages from all larvae cumulated). Experiments were performed in 2 replicates (cumulated in the graphs), each with 16–17 larvae per group. Total number of larvae analysed in C: 32 (cxcr4b+/+), 33 (cxcr4b−/−). Total number of macrophages analysed in D: 387 (cxcr4b+/+), 334 (cxcr4b−/−). (E–G) % of mpeg1:mCherry-F signal overlapping with Mm-GFP signal in cxcr4b−/− and cxcr4b+/+. No significant deviation in macrophage composition of the trunk granuloma lesions was detected. Each data point represents 1 granuloma. Experiment was performed in 1 replicate. Total number of granuloma analysed: 37 (cxcr4b+/+), 23 (cxcr4b−/−). Representative example images are shown in (F and G). (H) Expression of cxcr4b in macrophages (mpeg1+) and neutrophils (mpx+). Both cell subsets express cxcr4b at significantly higher levels than the negative (non-fluorescent) cell population. Data represent fold changes relative to the negative cell fraction. Cells were sorted at 2 dpf, from at least 100 embryos. Experiment was performed in 3 replicates. I-J. Macrophages are indispensable to mediate granuloma vascularisation (I) and an increased number of neutrophils (by irf8 morpholino knockdown) cannot compensate for macrophage deficiency, neither in cxcr4b−/− nor in cxcr4b+/+. Deficient bacterial expansion in irf8 knockdown condition (irf8-MO) compared to standard control morpholino treatment (Stctr-MO) can be attributed to the lack of vascularisation as suggested by non-significant differences in burden between the knockdowns and control cxcr4b−/− (J). Data are analysed as in Fig. 2A,B, but infections were performed at 33 hpf, instead of 2 dpf. Experiments were performed in 1 replicate. Total number of granuloma analysed: 47 (Stctr-MO-cxcr4b+/+), 47 (Stctr-MO-cxcr4b−/−), 61 (irf8-MO-cxcr4b+/+), 73 (irf8-MO-cxcr4b−/−). |
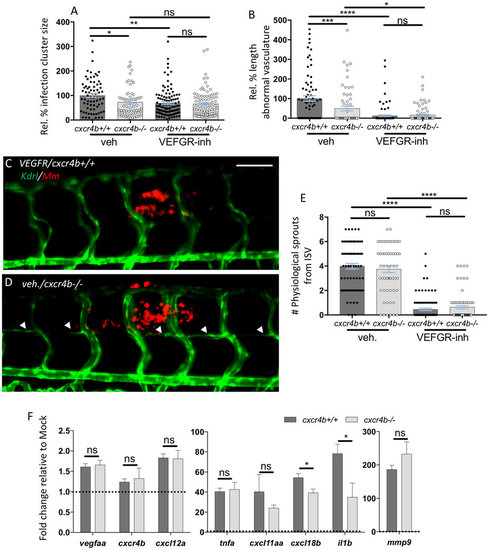
Deficient granuloma vascularisation in cxcr4b mutation does not strictly depend on aberrant Vegf signalling or attenuated expression of inflammatory genes. (A,B) cxcr4b deficiency reduced granuloma expansion to a similar extent as blockade of Vegf signalling (VEGFR-inh, treatment with Sunitinib) (A), although Vegf inhibition can more severely affect angiogenesis. (B). Data are analysed as in Fig. 2A,B. Experiments were performed in 2 replicates (cumulated in the graphs). Total number of granulomas analysed: 73 (vehicle-cxcr4b+/+), 66 (vehicle-cxcr4b−/−), 109 (VEGFR-inhibitor-cxcr4b+/+), 88 (VEGFR-inhibitor-cxcr4b−/−). C–E Inhibition of Vegf signalling (C), but not depletion of cxcr4b (D), severely affects physiological angiogenesis. Quantification in E was performed by counting the number of sprouts from intersegmental vessels (ISV) from images encompassing 6–7 somites in the trunk region (as represented in C,D). Experiment was performed in 2 replicates (cumulated in the graph). Total number of region of interest analysed: 73 (vehicle-cxcr4b+/+), 59 (vehicle-cxcr4b−/−), 93 (VEGFR-inhibitor-cxcr4b+/+), 77 (VEGFR-inhibitor-cxcr4b−/−). Scale bar: 100 μm. (F) At 5 dpi, Cxcr4b does not exert a transcriptional control on vegfaa expression, but affects the inflammatory response to mycobacterial infection, in particular by dampening il1b expression. qRT-PCR were performed whole mount from pools of at least 10 embryos (4 replicates). Data represent fold changes of Mm infection groups relative to their mock injection (2% PVP in PBS) control group (set to 1, dotted line). No significant differences in basal expression levels were found between mutants and wt for the analysed genes. |
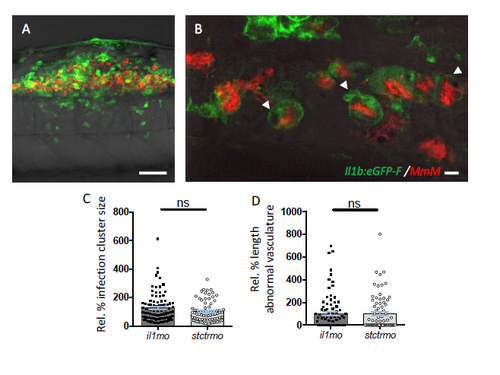
Function of il1b on trunk granulomas. A-B. Induction of the proangiogenetic inflammatory mediator il1b at the trunk granuloma. The large granuloma forming in the trunk contains a high number of il1b-expressing cells, predominantly consisting of infected cells. Confocal images were acquired at 5 dpi from infected Tg(il1b:eGFP-F) larvae. Scale bars: A: 50 μm; B: 5 μm. C-D. Despite being largely induced in infected cells during trunk granuloma formation, and notably suppressed by cxcr4b mutation, depletion of il1b does not recapitulate cxcr4b phenotype in granuloma vascularisation, since il1b morphants could still develop normally vascularise granulomas. Data were obtained and analysed as in Figure 3I-J (2 replicates, cumulated). |