
- Title
-
E-cadherin is required for gastrulation cell movements in zebrafish
- Authors
- Shimizu, T., Yabe, T., Muraoka, O., Yonemura, S., Aramaki, S., Hatta, K., Bae, Y.K., Nojima, H., and Hibi, M.
- Source
- Full text @ Mech. Dev.
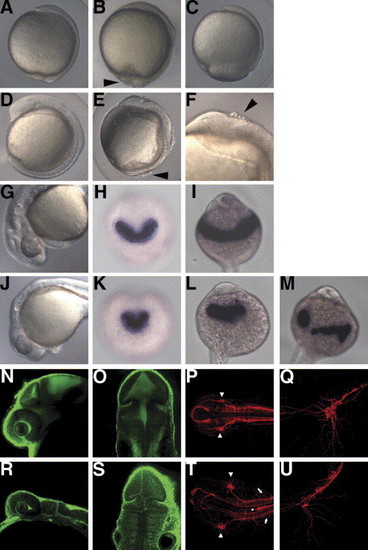
The rk3 mutant shows cell detachment, epiboly delay, and abnormalities in the formation of neural tissues and the hatching gland. (A–C) Morphology of the wild-type (A) and rk3 homozygous mutant embryos (B, C) at 10 hpf (bud stage for wild-type embryos). Lateral views with dorsal to the right. A group of the embryos completed epiboly at 10 hpf, but showed an irregular embryonic surface, distorted dorsal midline, and dissociation of cells (B, arrowhead). Severely affected embryos showed a delay in epiboly (C). The severity of the mutant phenotypes varied depending on the mating pairs and culture temperature (Fig. 2). (D–F) Morphology of the wild-type (D) and rk3 mutant embryos (E, F) at the segmentation stage (10-somite stage). (F) High magnification of the anterior region of the mutant embryo. The mutant embryos had a shorter anterior–posterior axis and wider somites than wild-type embryos, and cells that were dissociated from the embryos (indicated by arrowheads in E and F). The embryos often underwent yolk lysis and died during the segmentation period. (G,J) Morphology of the wild-type (G) and rk3 mutant embryos (J) at the pharyngula stage (24 hpf). Anterior is to the left. (I, L). In the mutant embryos, the anterior neural tissue was flattened and the hatching gland stayed in the midline and did not expand laterally. (H, I, K–M) Expression of the hatching gland marker hgg1 in the wild-type (H, I) and the rk3 mutant embryos (K–M) at the early segmentation stage (H, K) and at 24 hpf (I, L, M). Anterior views. The polster was wide in the medio-lateral axis in the wild-type embryos (H), but stayed in the midline at the segmentation stage in the rk3 mutant embryos (K). The hatching gland stayed on the front of the head (L), or was broken apart and mislocated (M) in the rk3 mutant embryos. (N, O, R, S) Bodipy-ceramide staining of the wild-type (N, O) and the rk3 mutant embryos (R, S) at 30 hpf. Confocal microscopic sagittal (N, R) and cross (O, S) sections. The rk3 mutant embryos showed a flattened nervous structure. (R, S, V, W) Immunostaining with anti-acetylated tublin at 30 hpf in wild-type (P, Q) and rk3 mutant (T, U) embryos. Dorsal views with anterior to the left (P, T). High-magnification views of the trigeminal ganglia and lateral views with anterior to the left (Q, U). The trigeminal ganglia (arrowheads in P, T) were located more laterally, and the neurons in the trigeminal ganglia were scattered or loosely organized, and their axons were disorganized in the rk3 mutant embryos. Abnormal positioning of Rohon-Beard neurons (arrows) and aberrant axon tracts in the dorsal midline (dot in T) were observed in the posterior hindbrain and the anterior spinal cord in the rk3 mutant embryos. EXPRESSION / LABELING:
PHENOTYPE:
|

The rk3 mutation is a temperature-sensitive allele. Embryos obtained from crossing five different pairs (1–5, D) of the rk3 heterozygotes were divided into two groups: one was incubated at 28.5 °C and the other at 34 °C. The phenotypes of the embryos were classified into four categories based on morphological inspection. Embryos that died by 24 hp were classified as ‘dead’. Class I embryos survived over 24 hpf, and had a very short or no tail structure and no discernible structure of the tectum and/or the midbrain–hindbrain boundary (MHB) (B). Class II embryos survived over 24 hpf, and had a tail and flattened neural structure with discernible midbrain and MHB structures (C). Embryos that did not show apparent abnormalities at 24 hpf were classified as ‘wild-type’ (wt) (A). The percentages of each class of embryos are indicated in the bar graph (D). Although the phenotypes of the rk3 embryos varied by parent pair, they were more severe in embryos incubated at 28.5 °C than at 34 °C. |
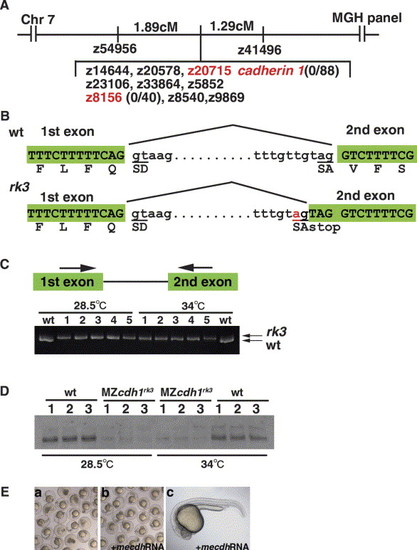
The rk3 locus encodes E-cadherin. (A) The rk3 locus was mapped to chromosome 7 close to the SSLP marker z8156 (no recombination in 40 meioses). The e-cadherin (cdh1) gene is located near z20715 in the radiation hybrid T51 panel, which was mapped adjacently with z8156 in the MGH panel. We found no recombination of a RFLP marker within the cdh1 gene in 88 meiotic events. (B) The rk3 allele contains a mutation in the first intron of the cdh1 gene. The mutation introduces an aberrant splicing acceptor and a premature stop codon on the 52 side of the codon of Val-71. (C) The rk3 locus generates an abnormal splicing variant. The rk3 zygotic homozygous embryos were cultured at 28.5 or 34 °C. RNAs from mutant embryos that survived more than 24 hpf were isolated and subjected to RT-PCR using primers that correspond to the 32 end of the first exon and the 52 end of the second exon. The PCR products were separated in a 20% polyacrylamide gel. The PCR product in each lane was derived from a single embryo. The mutant embryos had a longer transcript irrespective of the culture temperature. (D) E-cadherin protein was severely decreased in the maternal-zygotic (MZ) cdh1rk3 mutants. Embryos obtained from crossing the homozygous cdh1rk3 mutants, which were rescued by the expression of mouse e-cadherin, were grown at 28.5 or 34 °C, until wild-type (wt) embryos born at the same time reached the five-to-six somite stage. The embryo lysates were separated by SDS-PAGE and immunoblotted with anti-zebrafish E-cadherin antibodies. (E) Injection of 400 pg mouse e-cadherin (mcdh1) RNA suppressed the phenotypes of the zygotic rk3 mutant phenotypes (Eb, c). Ea, control. |
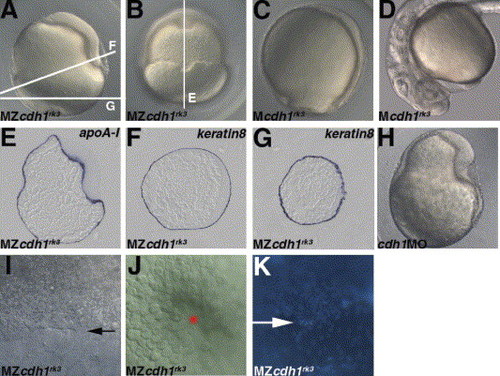
Maternal-zygotic (MZ) cdh1rk3 mutant embryos show epiboly arrest. (A, B) MZcdh1rk3 mutant embryos displayed epiboly arrest at 10 hpf. MZcdh1rk3 mutant embryos were obtained by crossing the homozygous cdh1rk3 fish, which were rescued by mcdh1 RNA injection. Lateral views with dorsal to the right (A). Dorsal views (B). (C, D) Maternal (M) cdh1rk3 mutant embryos displayed a mild epiboly delay at 10 hpf (C) and normal anterior neural tissues and hatching glands (D). Mcdh1rk3 mutant embryos were obtained by crossing the homozygous cdh1rk3 mutant fish and wild-type fish. Lateral views with dorsal to the right (C). Lateral views in the anterior region (D). (E–G) Expression of the yolk syncytial marker apoA-I (E) and enveloping layer marker keratin8 (F, G) at 10 hpf in the MZcdh1rk3 mutant embryos. The stained MZcdh1rk3 mutant embryos were sectioned as indicated by the white lines in A and B. (H) Morphology of the embryos that received 50 pg of cdh1MO (recognizing the splicing donor of the first intron) at 10 hpf. The cdh1 morphant embryos showed a phenotype similar to that of MZcdh1rk3 mutant embryos. (I, J) DIC images of the margin of the deep cells (I) (marked by an arrow) and enveloping layer (J) at the vegetal pole (marked by a star) in the MZcdh1rk3 mutant embryos at 10 hpf. Lateral views (I) and vegetal pole views (J). The enveloping layer but not the deep cells reached the vegetal pole. (K) DAPI staining of 10 hpf MZcdh1rk3 mutant embryos. Dorsal views. The margin of the deep cells is indicated by an arrow. On the animal pole side, there was a high density of DAPI-stained nuclei, indicating the deep cells. On the vegetal pole side, the nuclei were distributed sparsely, indicating the yolk syncytial layer. |
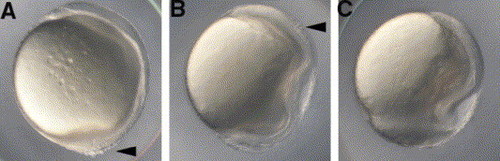
The cdh1rk3 allele does not complement weg. Embryos obtained from crossing weg male fish with cdh1rk3 female fish showed abnormal morphology at 10.5 hpf; (A) relatively normal morphology with dissociation of cells (arrowhead), (B) irregular cell surface and distorted dorsal axis with dissociation of cells (arrowhead), and (C) epiboly arrest. Lateral views, with dorsal to the right. The numbers of embryos that displayed each phenotype were (A) 6, (B) 42, (C) 9, and normal morphology 110 (n=167). |
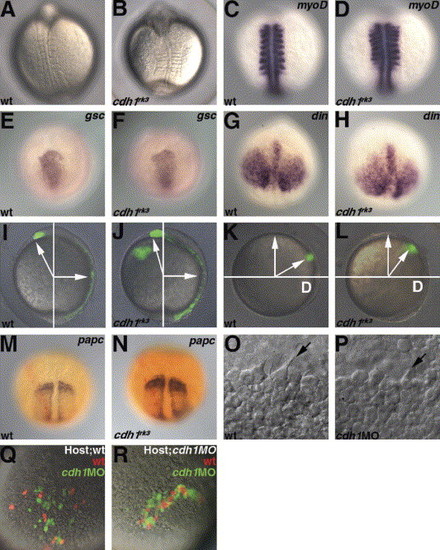
Impaired gastrulation cell movements in the cdh1rk3 mutants. (A, B) Dorsal views of the wild-type and the zygotic cdh1rk3 mutant embryos at the ight-somite stage. (C, D) Expression of myoD in the wild-type and the cdh1rk3 mutant embryos at the eight-somite stage. Dorsal views. (E–H) Expression of goosecoid (gsc) and chordin (din) in the wild-type (E, G) and the cdh1rk3 mutant embryos at 8.5 hpf. Animal pole views with dorsal to the bottom (E, F) and dorsal views with anterior to the top. (I–L) Impaired convergence and extension. Cells in the embryonic shield (I, J) or cells at the lateral blastoderm margin (90 °C from the dorsal side, K, L) in wild-type (I, K) and cdh1rk3 mutant (J, L) embryos were labeled by uncaging fluorescein dextran with ultraviolet light at the shield stage. The position of the labeled cells was determined at 10 hpf. Lateral views with dorsal to the right (I, J), and vegetal pole views with dorsal to the right (K, L). The angles for the anterior movements (extension and migration) of the anterior mesendoderm were 121.8±4.3° (average±SD, n=8) in the wild-type and 110.8±4.4° (n=5) in the cdh1rk3 mutant embryos. The angles for the convergence movements of the lateral blastoderm margin were 59.4±5.9° (n=7) in the wild-type and 50.7±11.2° (n=7) in the cdh1rk3 mutant embryos. Embryos in which the lateral blastoderm margin was labeled were fixed at 10.5 hpf (the two-somite stage for wild-type) and stained with papc riboprobe (purple) and anti-fluorescein antibodies (red) (M, wild-type; N, cdh1rk3 mutant). (O, P) DIC images of the anterior axial mesendoderm in the wild-type (O) and cdh1 morphant mutant embryos (P) at 8.5 hpf (80% epiboly for the wild-type). The filopodia formation (arrows) of the anterior axial mesendoderm was observed in both the wild-type and the cdh1 morphant embryos. (Q, R) Transplantation of wild-type and Cdh1-defective cells into wild-type (Q) and cdh1 morphant embryos (R). Cells in the blastoderm margin were isolated from wild-type control (marked by Alexa Fluor 568 dextran, red) and cdh1 morphant (marked by fluorescein dextran, green) embryos and transplanted into the ventral margin of shield stage wild-type embryos. The locations of the control and the cdh1 morphant cells were determined at 9 hpf. Both the transplantation experiments were carried out nine times and the representative data were shown. Ventro-posterior (Q) and ventral (R) views. EXPRESSION / LABELING:
PHENOTYPE:
|
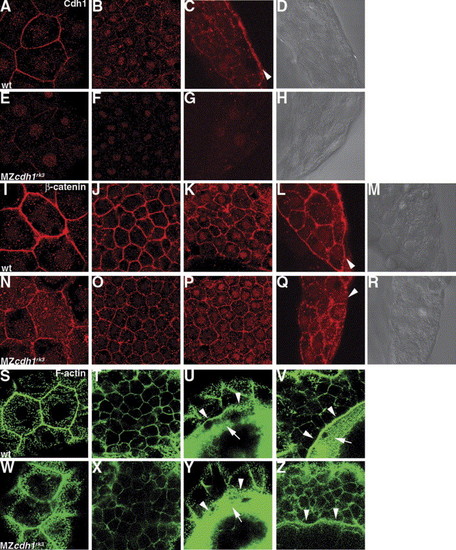
Cell adhesion within DCs and microfilament formation at the margin of the EVL and DCs are maintained in the cdh1 morphant embryos. Wild-type (wt) and MZcdh1rk3 mutant embryos were fixed at 8.5 hpf (80% epiboly for wild-type), and stained with an anti-Cdh1 antibody (A–H), anti-β-catenin antibody (I–R), or Alexa Fluor 488-phalloidin (S–Z). Immunostaining was visualized by Alexa Fluor 568-conjugated secondary antibody (A–R). (A–D, I–M, S–V) Wild-type embryos. (E–H, N–R, W–Z) MZcdh1rk3 mutant embryos. Confocal microscopic optical sections in the animal pole region at the level of the enveloping layer (A, E, I, N, S, W) and deep cells (B, F, J, O, T, X). Confocal sections in the lateral blastoderm margin (C, D, G, H, K–M, P–R), cross sections of the dissected embryos (C, D, G, H, L, M, Q, R), and optical sections at the level of the deep cells (K, P). (D, H, M, R) DIC images of (C, G, L, Q). Confocal optical section of the epiboly margin at the level of the EVL (U, Y) and deep cells (V, Z). Cdh1 protein was strongly detected in the contact surface between the EVL and the DCs (arrowhead in C). Accumulation of β-catenin was detected on the contact surface between the YSL and the DCs in the wild-type embryos but was severely decreased in the MZcdh1rk3 mutant embryos (arrowheads in L, Q). The ring-like F-actin-based structures were detected in the edges of the EVL (arrowheads, U, Y) and the edge of the DCs (arrowheads in V, Z), and punctate actin bands were detected in the E-YSL (arrows in U, V, Y), adjacent to the edges of the EVL and the DCs in the wild-type embryos, and to the edge of the EVL in the MZcdh1rk3 mutant embryos. EXPRESSION / LABELING:
|
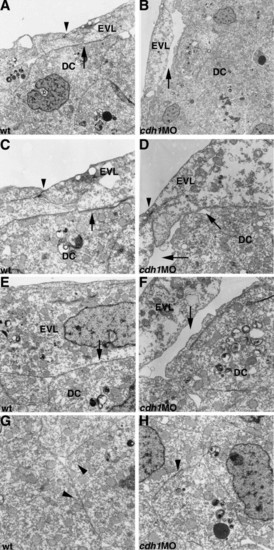
Cell adhesion between the deep cells and the enveloping layer is disrupted in cdh1 morphant embryos. Wild-type control (wt, A, C, E, G) and cdh1 morphant embryos (MO) embryos (B, D, F, H) were fixed at 8.5 hpf (80% epiboly for the wild-type) and subjected to electron microscopic analysis. Lateral regions. Adhesion between the enveloping layer (EVL) and the deep cells (DCs) was detected in the wild-type control embryos (indicated by arrows in A, C, E), but was severely affected in the cdh1 morphant embryos (arrows in B, D, F). In contrast, the adhesions between the EVL cells (arrowheads in A, C, D) or between the DCs (arrowheads in G, H) were not significantly affected in the cdh1 morphant embryos, compared with the wild-type control embryos. PHENOTYPE:
|
Reprinted from Mechanisms of Development, 122(6), Shimizu, T., Yabe, T., Muraoka, O., Yonemura, S., Aramaki, S., Hatta, K., Bae, Y.K., Nojima, H., and Hibi, M., E-cadherin is required for gastrulation cell movements in zebrafish, 747-763, Copyright (2005) with permission from Elsevier. Full text @ Mech. Dev.