
- Title
-
Splitting of circulating red blood cells as an in vivo mechanism of erythrocyte maturation in developing zebrafish, chick and mouse embryos.
- Authors
- Brönnimann, D., Annese, T., Gorr, T.A., Djonov, V.
- Source
- Full text @ J. Exp. Biol.

In vivo proliferation and splitting of circulating red blood cells (RBCs) in zebrafish embryos. (A) Overview of a zebrafish embryo at 48 hours post-fertilization (hpf) in vivo. ISVs, intersegmental vessels; CVP, caudal vein plexus. (B) Phospho-histone H3 (pH3) staining (blue) of the zebrafish caudal vein plexus. In Tg(fli1a:GFP;gata1:DsRed) double transgenic animals, endothelial cells (green) and RBCs (red) are labeled through expression of fluorescent markers as indicated. (C) Example of a pH3-positive circulating RBC. (D) Average extent of pH3-positive cells as percentage of circulating gata1-positive RBCs (n=9; >100 quantified RBCs per animal). (E) Splitting of proliferating RBCs is a four-step process. Following cell division (i), the two somata of a dividing RBC might eventually tangle (ii) at a vascular bifurcation which leads to the subsequent elongation (iii) of the soma-to-soma connection and its eventual rupture (iv). (F) We observed 11.24±0.93 RBC splitting events per hour (n=11; 142 events in total). (G) On average, a splitting event (i.e. phase ii–iv) occurred within 129.56±1.29 s (n=11; 142 events). |
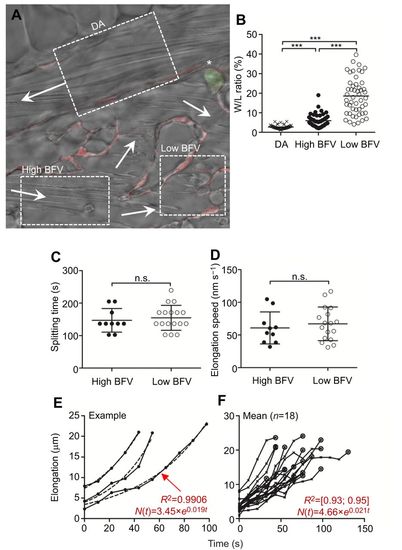
RBC splitting is independent of the incident blood flow velocity in zebrafish embryos in vivo. To assess the impact of blood flow velocity (BFV) as a major shear force determinant on splitting time and elongation speed during RBC splitting, we compared RBC splitting at two regions receiving either high or low BFV. (A) Three regions in a Tg(fli1a:RedX;mpx:eGFP) fish, the dorsal aorta (DA), high BFV and low BFV, are indicated. Arrows mark the respective blood flow direction. The asterisk indicates a primitive neutrophil. (B) BFV was deduced by the width/length (W/L) ratio of adjacent circulating blood cells. Images were acquired throughout a constant period of time. In regions with high BFV (i.e. DA), RBCs are elongated (i.e. L>W, low W/L ratio), while at low BFV, RBCs appear more rounded (i.e. increased W/L ratio). Therefore, the W/L ratio serves as a proxy for the local BFV. ***P<0.001. (C) Splitting time and (D) elongation speed were both independent on BFV. (E,F) Exponential growth (E) and elongation length (F). Once RBCs are tangled at vascular structures (phase ii), the soma-to-soma connection subsequently elongates (phase iii). The process of elongation can be mathematically expressed using exponential growth equations. This implies that the longer a cell is tangled, the faster its retained connection elongates. On average, elongation was represented by the equation N(t)=4.66×e0.021t with a regression coefficient (R2) between 0.93 and 0.95 (n=1 animal, n=18 events). |
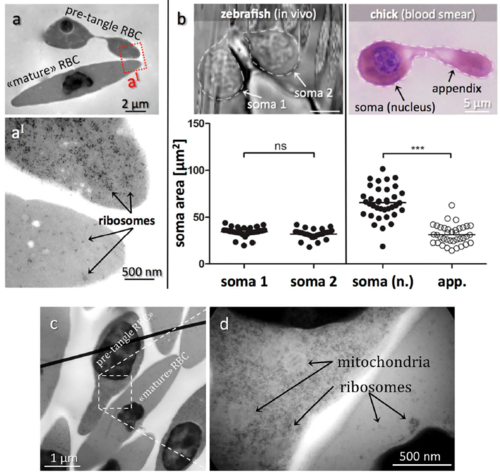
Transmission electron micrographs of RBCs in blood of fish and chick embryos. (a) Electron micrographs comparing pre-tangle and mature chick RBCs. (a´) Zoomed region indicated in (a) in a pre-tangle RBC (top) contains abundant free ribosomes in contrast to the mature RBC (bottom). Other side-by-side comparisons of pre-tangling versus mature chick RBCs as shown in (c + d) confirmed the far higher quantity of ribosomes and mitochondria in immature RBCs relative to fully differentiated cells. (b) Comparing somata sizes of dumbbellshaped zebrafish and chick RBCs. In the zebrafish, both somata are nucleated and of similar size. In contrast, only one soma is nucleated in the chick RBC, which is significantly larger in size than the other soma. ***P<0.001. |