
- Title
-
Inhibitors of neutrophil recruitment identified using transgenic zebrafish to screen a natural product library
- Authors
- Wang, X., Robertson, A.L., Li, J., Chai, R.J., Haishan, W., Sadiku, P., Ogryzko, N.V., Everett, M., Yoganathan, K., Luo, H.R., Renshaw, S.A., and Ingham, P.W.
- Source
- Full text @ Dis. Model. Mech.
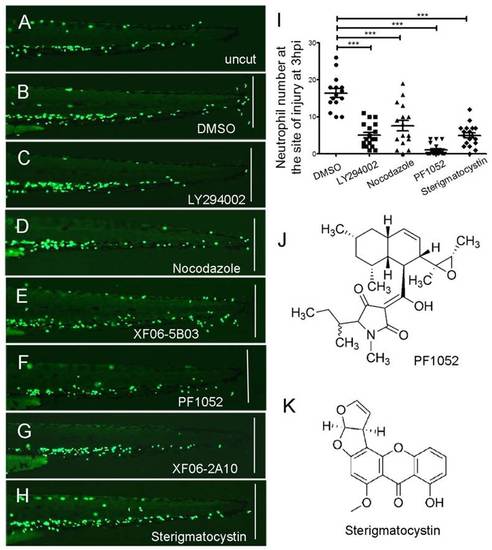
Inhibitory effect of PF1052 and sterigmatocystin on neutrophil migration towards the wound site. Neutrophil migration assay was carried out using Tg(mpx:GFP)i114 larvae according to Renshaw et al. Renshaw et al., 2006). (A) Neutrophils were quiescent on an uninjured 3-dpf larva. (B) About ten neutrophils were recruited to the wound site 3 hours after the tailfin was amputated on a control larva that was treated with DMSO. (C,D) Larvae treated by PI3K inhibitor LY294002 (50 µM) or microtubule inhibitor nocodazole (33 µM), respectively, had few neutrophils recruited to the wound. (E) An extract (ID: XF06-5B03) from an ascomycete genus Sphaeropsidales completely blocked neutrophil migration towards the wound at 50 µg/ml. (F) The active component identified from XF06-5B03 extract (PF1052) also completely blocked neutrophils recruitment at a very low concentration of 2 µM; the molecular structure of PF1052 is shown in J. (G,H,K) Another extract, XF06-2A10, from a fungus, genus Aspergillus, that blocked the migration of neutrophils contains the functional component sterigmatocystin, working at 50 µM. (I) Quantification of neutrophils recruited to the site of injury (n>15). The white line on B-H indicates the amputation site of tailfin. ***P<0.001. |
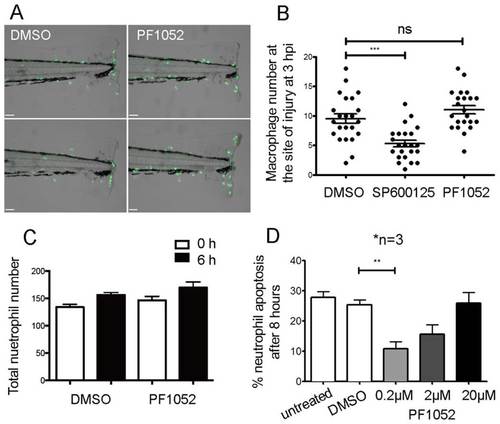
PF1052 is a specific inhibitor of neutrophil migration. (A,B) PF1052 has no significant effect on macrophage migration; the MAP kinase inhibitor SP600125 was used at 30 µM as a positive control. (C) PF1052 did not significantly reduce total neutrophil number in intact larvae. (D) PF1052 does not induce human neutrophil apoptosis and seems to promote survival at a low concentration (**P<0.01; ***P<0.001; ns, not significant). |
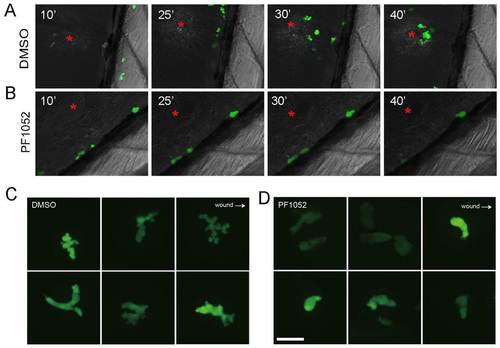
Time-lapse imaging of neutrophil migration in vivo. The ventral fin of 6-dpf larvae was injured by a sharp syringe needle, and chemotaxis of neutrophils were imaged by spinning disk microscope. (A) In a control larva treated with DMSO, neutrophil migration was not directed until 10 minutes after fin injury. Thereafter, they migrated actively towards the wound site (red asterisk; 25 and 30 minutes); by 40 minutes after injury, around eight neutrophils were recruited. (B) In contrast to the control, neutrophils treated with PF1052 remained stationary during 40 minutes of imaging; they appeared round and barely formed any pseudopods. (C) Higher magnification shows that DMSO-treated neutrophils actively formed multiple pseudopods and this was impaired in PF1052 treated-neutrophils (D). |
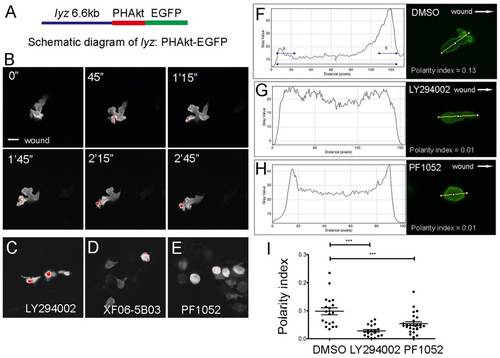
PF1052 inhibited neutrophil migration in a PI3K-independent manner. (A) Schematic diagram of the lyz:PHAkt-EGFP construct used to generate the stable transgenic line. (B) Time-lapse imaging of migrating neutrophils showed PHAkt-EGFP signal was always localised to the cell front in control larvae. The red signal indicates the highest level of PI3K at the leading edge. (C) PI3K inhibitor LY294002 reversed the cellular localisation of PI3K, which accumulated within the cell body. (D,E) Extracts from Sphaeropsidales and its functional component PF1052 did not affect the localisation of PI3K at the cell membrane. (F-I) Quantification of neutrophil polarity index in DMSO-, LY294002- and PF1052-treated larvae. LY294002 and PF1052 significantly reduced neutrophil polarity index (***P<0.001). |