|
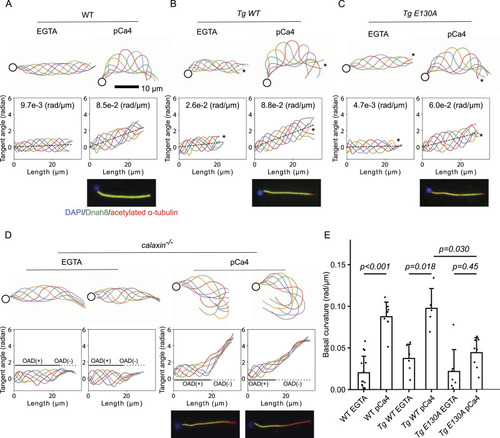
Calaxin calcium-binding activity is necessary for the calcium-induced asymmetric beating of sperm. (A, B, C, D) Traces (top) and tangent angle plots (bottom) of WT (A), Tg WT (B), Tg E130A (C), and calaxin−/− (D) demembranated sperm models under the presence of 0.1 mM EGTA (EGTA) or CaCl2 (pCa4). Immunofluorescence microscopy results from Fig 1C are also shown for the comparison. WT and Tg WT showed an almost symmetric waveform in the EGTA condition, and the pCa4 condition induced asymmetric waveform. Tg E130A showed a similar waveform in the EGTA condition, but the pCa4 condition induced weaker asymmetry compared with WT and Tg WT. calaxin−/− sperm model had an active proximal region and an inactive distal region. The inactive distal region exhibited a highly curved waveform in the pCa4 condition. (A, B, C) Calculated basal curvatures are shown in tangent angle plots. (B, C) Asterisks indicate distal ends of flagella that lack an outer arm dynein (OAD). (D) Black lines and dotted lines show estimated OAD(+) and OAD(−) regions, respectively. (E) Comparison of basal curvatures of WT, Tg WT, and Tg E130A sperm flagella. The pCa4 condition induced significantly higher basal curvature in both WT and Tg WT, but Tg E130A did not show a significant elevation of basal curvature. In addition, the basal curvature of Tg E130A was significantly lower than that of Tg WT in the pCa4 condition. All data are shown with a mean (bar graphs) + SE (error bars). All data were collected from n = 14 (WT EGTA), 8 (WT, pCa4), 6 (Tg WT EGTA), 5 (Tg WT pCa4), 7 (Tg E130A EGTA), and 8 (Tg E130A pCa4) sperm flagella from two or more independent experiments.
|