
- Title
-
Regulation of nodal signalling and mesendoderm formation by TARAM-A, a TGFbeta-related type I receptor
- Authors
- Aoki, T.O., Mathieu, J., Saint-Etienne, L., Rebagliati, M.R., Peyriéras, N., and Rosa, F.M.
- Source
- Full text @ Dev. Biol.
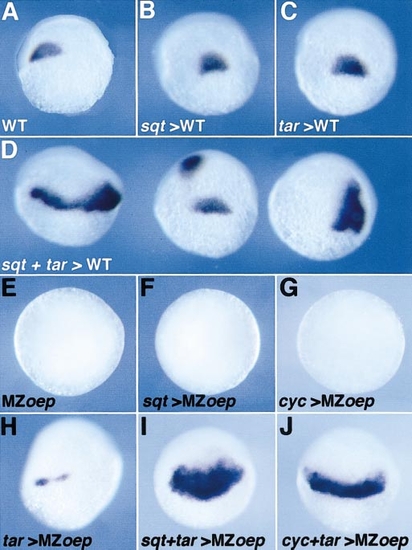
Functional interaction between sqt, cyc, and tar. (A–D,shield stage, dorsal view) (A) Expression of gsc in control gastrulae or (B) in gastrulae overexpressing sqt RNA (0.2 pg), (C) tar RNA (100 pg), or (D) both as indicated. Note the enlargement or the duplication of the gsc domain in (D). (E–J) A similar overexpression of sqt, cyc, tar RNAs, or a combination as indicated was carried out in MZoepm134 embryos and the expression of gsc was analysed when embryos had reached the early gastrula period (equivalent to the shield stage). (E–G, animal pole view) gsc is not expressed in control uninjected embryos or embryos overexpressing sqt, or cyc RNAs. (H–J, lateral view) tar overexpression induces a weak expression of gsc in some embryos (H) but strongly induces the ectopic expression of gsc when coexpressed with cyc or sqt (I, J). |
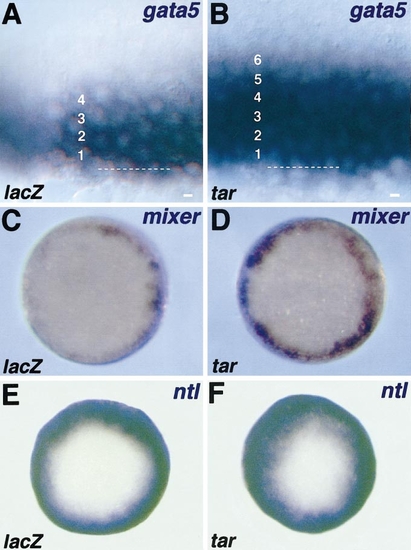
tar overexpression expands the expression domains of early mesendodermal markers. Embryos were injected with lacZ RNA (200 pg) as a control or with tar RNA (200 pg), fixed during the late blastula period (40% epiboly), and analysed by in situ hybridisation with the marker indicated (upper right). (A, B, lateral view) At this stage, gata5 is normally expressed (A) in four rows of cells at the margin. (B) In tar mRNA-injected embryos, the expression of gata5 is expanded to the fifth and sixth rows. The dotted line indicates the YSL-blastoderm border. (C, D, animal pole view) Similarly, the normal expression of mixer (C) encompasses three to four rows of marginal cells with a patchy appearance. (D) The overexpression of tar leads to an expansion of the mixer expression domain, which appears more uniform. (E, F, animal pole view) The pan-mesodermal marker ntl is expressed in larger domain than gata5 at the margin of control embryo encompassing 8–10 cell diameters (E, not shown). When tar is overexpressed, the ntl expression is expanded to animal pole (F). |
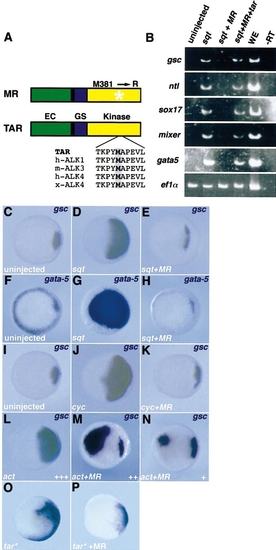
TarMR interferes with the inducing activities of cyc and sqt. (A) Structures of Tar MR381 (MR). Wild-type Tar (TAR) is composed of an extracellular domain (EC), a transmembrane domain (black), aGS domain (GS), and a kinase domain (Kinase). TarMR (MR) carries a point mutation that substitutes an arginine for methionine 381 in the kinase domain (star). This methionine is highly conserved between zebrafish TAR, human ALK1, mouse ALK3, human ALK4, and Xenopus ALK4 (boxed). (B) Embryos were injected with sqt RNA (2 pg), combined with tarMR RNA (20 pg) alone or with tarRNA(100 pg), the animal caps were dissected, cultivated until the shield stage, and analysed by RT-PCR for different mesendodermal markers. tarMR blocks the effect of exogenous nodal-related molecules on meso- or endodermal markers as indicated. Ef1α is used as a loading control. In the -RT lane, the reverse transcription step has been omitted. In the WE lane, RT-PCR products from whole embryos were loaded. (C–P) Embryos were injected with sqt, cyc, activin (act), tarMR, tar*, RNA, or a combination as indicated, fixed at the shield stage, and analysed by in situ hybridisation with the marker indicated in the upper right corner. Animal pole views, dorsal to the right when it can be determined. |

tarMR interferes with the formation of anterior midline structures. Wild-type embryos were either left uninjected or injected with tarMR RNA (MR) alone or with wild-type tar RNA (MR + tar) at the 16-cell stage into one marginal blastomere. The phenotypes of the embryos were analysed at 24–30 hpf, either on live embryos (A–C, frontal head view) or by in situ hybridisation (D–F, lateral view, anterior to the left) with the ventral CNS marker shh. (A–C) Overexpression of tarMR leads to cyclopia in 11% (n = 95) of tarMR-injected embryos (B) which can be rescued (C, remaining cyclopic embryos 3%, n = 75) by coexpression of wild-type tar RNA. (D–F) Similarly, overexpression of tarMR leads to the downregulation of shh in 12% of tarMR-overexpressing embryos in the ventral brain (E, n = 95). This phenotype was rescued by adding 100 pg of tar RNA (F). |
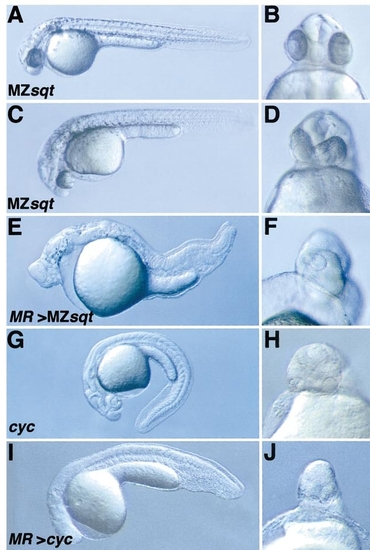
tarMR enhances the phenotype of mutants with attenuated Nodal signalling. tarMR RNA was injected during the first cell cycle into mutant embryos and phenotypes of the injected embryos were analysed on 24- to 30-hpf live embryos presented as a lateral view from whole embryos (left column, anterior to the left) or as a frontal head view (right column). (A–D) Phenotypes of MZsqt mutants. The cross between homozygous individuals results in maternal–zygotic sqtcz35 mutants (MZsqtcz35) which exhibit a variable phenotype from almost normal (A, B) to more severe ones (C, D). When tarMR is overexpressed into MZsqtcz35 embryos, their phenotype is dramatically enhanced (E, F, 100%, n = 65). (G, H) Phenotypes of cycb16 homozygotes. Uninjected embryos exhibit a mild cyclopia and the absence of floor plate. When tarMR is overexpressed, their phenotype is slightly enhanced (I, J). |
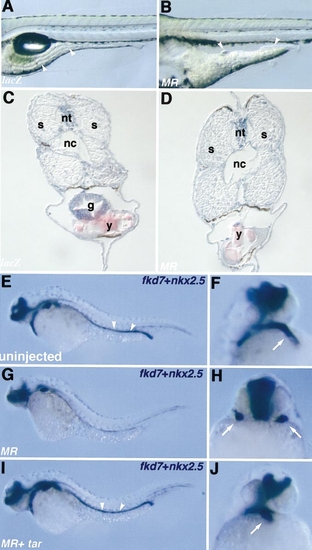
tarMR overexpression disrupts the formation of endoderm-derived structures and heart organogenesis. Wild-type embryos were either left uninjected or injected during the first cell cycle with lacZ RNA (200 pg), tarMR RNA (MR) (200 pg) alone, or combined with tar RNA (100 pg) as indicated in the lower left corner, analysed at 4 days as live samples (A, B), then fixed and sectioned (C, D) or at 30 hpf by in situ hybridisation (E–J). (A–D) At 4 days, large segments of the gut (arrowheads) are missing in em>tarMR-injected embryos (B, D) as compared to controls (A, C). (E) At 30 hpf, fkd7 labels the digestive structures from the pharynx to the digestive tube (arrowheads), as well as the ventral CNS. (G) Fkd7 expression is missing in digestive structures of tarMR expressing embryos but is restored upon coexpression of tar (I). (F, H, J) The same embryos were also labelled with the heart-specific probe nkx2.5 (arrow). A lateral view (F, J) or frontal view (H) is presented. Expression of tarMR leads to cardia bifida (H), which is rescued by coexpression of tar (J). Abbreviations: s, somite; nt, neural tube; nc, notochord; g, gut; y, yolk. |
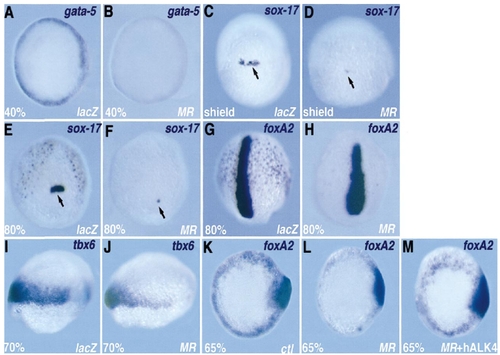
tarMR disrupts early endodermal marker expression. Embryos were injected with lacZ RNA (400 pg) as a control or with tarMR RNA (400 pg), fixed at the stage indicated in the lower left corner, and analysed by in situ hybridisation with the marker indicated (upper right). (A, B, animal pole view) gata5 is expressed in the YSL (not visible) and in the blastoderm. Blastodermal expression is lost in (B). (C–F, dorsal view) sox17 labels deep flat endodermal cells and a group of noninvoluting forerunner cells (arrow), which are severely reduced in number in (D) and (F) as compared to controls. (G, H, dorsal view) axial/foxA2 labels the nascent axis and deep scattered endodermal cells (arrowheads). In (H), the axis is unaffected but endodermal cells are missing. At 70% epiboly, tbx6 marks the lateral mesoderm, the precursors of the somites (I). This expression is very partially affected on the dorsolateral side (J). (K–M) The inhibition of endodermal staining of foxA2 by tarMR was rescued by coinjecting with human ALK4. |
Reprinted from Developmental Biology, 241(2), Aoki, T.O., Mathieu, J., Saint-Etienne, L., Rebagliati, M.R., Peyriéras, N., and Rosa, F.M., Regulation of nodal signalling and mesendoderm formation by TARAM-A, a TGFbeta-related type I receptor, 273-288, Copyright (2002) with permission from Elsevier. Full text @ Dev. Biol.