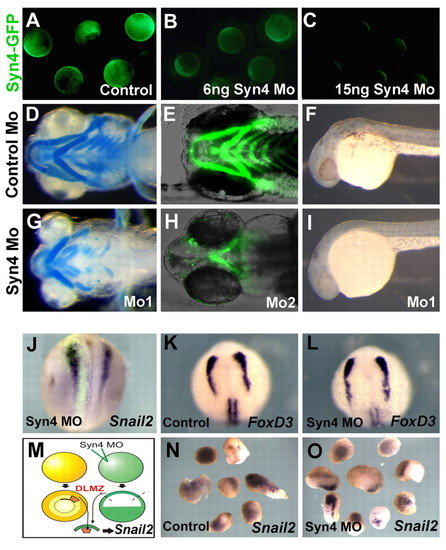
Syn4 is required for NC development but not for NC induction. (A-C) Efficiency of the syn4 MO. Zebrafish embryos were injected with 250 pg of syn4-egfp mRNA containing the MO-binding site (A). Fluorescence was significantly decreased by co-injection of 6 ng syn4 MO (B) and was totally abolished with 15 ng MO (C), demonstrating that the MO is able to efficiently inhibit Syn4. (D,E,G,H) Ventral view of zebrafish embryos showing cartilage development, analyzed by Alcian Blue staining (D,G) or cartilage fluorescence in the sox10:egfp line (E,H). (D,E) Control MO; (G) 6 ng of syn4 MO1; (H) 6 ng of syn4 MO2. (F,I) Lateral view of zebrafish embryos showing melanocytes. (F) Control MO; (I) syn4 MO. (J-O) Syn4 is not required for NC induction. (J) syn4 MO was co-injected with FDX into one blastomere of a two-cell stage Xenopus embryo and the expression of snail2 was analyzed at stage 18, before NC migration. No effect on the expression of snail2 was observed (96%, n=42). (K,L) Zebrafish embryos were injected with control MO (K) or syn4 MO (L), and the expression of the premigratory NC marker foxd3 was analyzed. No effect on the expression of foxd3 was produced by injection of syn4 MO. (M) syn4 MO was injected into one-cell stage Xenopus embryos, and at the blastula stage animal caps were dissected and conjugated with dorsolateral marginal zone (DLMZ), which is know to induce NC. The expression of snail2 was analyzed at the equivalent of stage 18. (N) Conjugates of DLMZ with control animal caps show a strong induction of snail2 (100%, n=15). (O) Conjugates of DLMZ with syn4-MO injected animal caps also show a strong induction of snail2 (100%, n=10).
|
