
- Title
-
Tmem2 restricts atrioventricular canal differentiation by regulating degradation of hyaluronic acid
- Authors
- Hernandez, L., Ryckebüsch, L., Wang, C., Ling, R., Yelon, D.
- Source
- Full text @ Dev. Dyn.
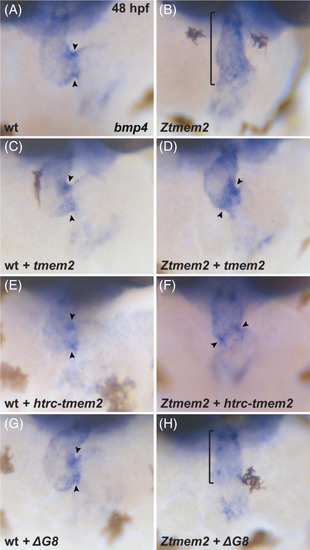
The ectodomain of Tmem2 is critical for proper AVC differentiation. In situ hybridization indicates expression of bmp4 in wild‐type (A, C, E, G) embryos and Ztmem2 (B, D, F, H) mutant siblings at 48 hours post fertilization (hpf); frontal views. In wild‐type (wt) embryos, concentrated expression of bmp4 is restricted to the AVC (A, arrowheads), whereas bmp4 is broadly expressed throughout the ventricle in Ztmem2 mutants (B, bracket). Expression of full‐length tmem2 can restore the enriched concentration of bmp4 expression in the AVC of Ztmem2 mutants (D, arrowheads; Table 1); additionally, expression of full‐length tmem2 does not affect bmp4 expression in wt siblings (C, arrowheads; Table 1). Expression of full‐length tmem2 also improves cardiac looping and AVC constriction in Ztmem2 mutants (D). Similarly, expression of htrc‐tmem2 can rescue the bmp4 expression pattern in Ztmem2 mutants (F, arrowheads; Table 1), but does not disrupt bmp4 expression in wt siblings (E; Table 1). Finally, as with ΔC‐term and all other extracellular domain deletion variants, expression of the ΔG8 variant provides no evident rescue of the bmp4 expression pattern in Ztmem2 mutants (H, bracket; Table 1) and does not affect bmp4 expression in wt embryos (G, arrowheads; Table 1) |

Subcellular localization of Tmem2 and Tmem2 variants in zebrafish blastomeres. Confocal reconstructions depict a lateral portion of the live zebrafish blastula at 4 hpf, following injection with mRNA encoding Tmem2‐GFP (A) or its variants (B‐I); the large size and optical accessibility of blastomeres are ideal for analysis of GFP localization. Full‐length Tmem2 and Htrc‐Tmem2 are both found at the plasma membrane (A, B). In contrast, Sc‐Tmem2 appears to traffic to the extracellular environment (C), while blastomere membranes remain intact (data not shown). All variants without extracellular domains lack evident membrane localization and, instead, display primarily cytoplasmic localization (D‐H). The R267H variant is also not apparent at the plasma membrane, yet its localization is distinct in that it is found in clusters within the cell (I). Although other variants are occasionally found in similar clusters (eg, H), they are especially prominent and frequently observed for R267H (I). Sample sizes were: tmem2, n = 12; htrc‐tmem2, n = 11; sc‐tmem2, n = 9; ΔC‐term, n = 10; ΔG8, n = 8; ΔPLT, n = 9; ΔPbH1, n = 10; ΔPL, n = 8; R267H, n = 16 |

The ectodomain of Tmem2 is critical for promoting cardiac fusion. In situ hybridization indicates expression of myl7 in Mtmem2 (A, D, G, I) and MZtmem2 (B, C, E, F, H, J) mutant embryos at 24 hpf; dorsal views, anterior up. The heart tube assembles normally in Mtmem2 embryos (A), with no evident defects or delay in cardiac fusion, just as is observed in wild‐type and Ztmem2 mutant embryos.12 MZtmem2 mutant siblings, however, display either cardia bifida (B) or, less frequently, partial cardiac fusion (C). Expression of full‐length tmem2 in MZtmem2 mutants can facilitate cardiomyocyte movement to the midline (E) and frequently restores heart tube formation (F) (Table 2). Expression of htrc‐tmem2 in MZtmem2 mutants can also rescue the MZtmem2 cardiac fusion defects (H; Table 2). As with ΔC‐term and all other extracellular domain deletion variants, expression of the ΔG8 variant does not rescue cardiac fusion in MZtmem2 mutants (J; Table 2). Mtmem2 sibling embryos appear unaffected by expression of tmem2 (D) or tmem2 variants (G, I; Table 2) |
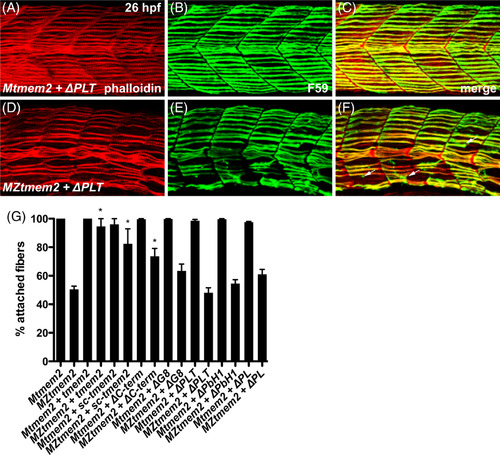
The ectodomain of Tmem2 is critical for promoting skeletal muscle fiber attachment. Immunofluorescence (A‐F) depicts muscle fiber organization, using phalloidin (red) to recognize both fast and slow muscle fibers and F59 (green) to recognize slow fibers; lateral views with dorsal up at 26 hpf. Somite morphology of Mtmem2 sibling embryos is indistinguishable from wild‐type, whereas MZtmem2 mutants display muscle fiber detachment.14 Although expression of full‐length tmem2 has been shown to rescue fiber detachment in MZtmem2 mutants,14 MZtmem2 mutants expressing ΔPLT still display detached fibers (D‐F; arrows in F mark examples of detached fibers). In contrast, Mtmem2 siblings expressing ΔPLT exhibit normal fiber attachment (A‐C). Bar graph (G) compares average prevalence of muscle fiber attachment at 26 hpf; error bars indicate SEM. In each case, we scored F59+ fibers in 10‐11 somites of the left myotome in multiple embryos. Only the F59+ fibers with intact attachments at both boundaries of the somite were scored as “attached”; fibers with dysmorphic attachment, partial detachment, or complete detachment at either end were scored as “detached,” including detached fibers that had retracted substantially. Asterisks indicate significant differences from uninjected MZtmem2 mutants, as determined by Student's t‐test. Full‐length tmem2 (P < .001), sc‐tmem2 (P = .0056), and ΔC‐term (P = .0014) provide significant rescue of muscle fiber attachment in MZtmem2 mutants, whereas other variants do not. Expression of tmem2 variants does not affect fiber attachment in Mtmem2 siblings. Graph includes previously published data for tmem2 and sc‐tmem2 (previously called “ectodomain”).14 Sample sizes were: Mtmem2, n = 4; MZtmem2, n = 7; Mtmem2 + tmem2, n = 6; MZtmem2 + tmem2, n = 8; Mtmem2 + sc‐tmem2, n = 2; MZtmem2 + sc‐tmem2, n = 5; Mtmem2 + ΔC‐term, n = 3; MZtmem2 + ΔC‐term, n = 5; Mtmem2 + ΔG8, n = 4; MZtmem2 + ΔG8, n = 5; Mtmem2 + ΔPLT, n = 4; MZtmem2 + ΔPLT, n = 7; Mtmem2 + ΔPbH1, n = 3; MZtmem2 + ΔPbH1, n = 4; Mtmem2 + ΔPL, n = 2; MZtmem2 + ΔPL, n = 5 PHENOTYPE:
|

Ztmem2 mutants exhibit excess HA deposition. Immunofluorescence with MF20 (red) to label the myocardium and HABP (green) to label HA in single optical slices through wt (A‐C) and Ztmem2 mutant (D‐F) hearts at 48 hpf. In wt, a thin layer of HA is visible throughout the ventricle, adjacent to the myocardium, and increased concentration of HA is visible within the AVC (A, arrowheads). In contrast, Ztmem2 mutants display increased levels of HA throughout the ventricle and at the AVC (D, arrowheads). Separate optical slices (G‐L), 2.5 microns away from those shown in (A‐F), provide a better view of the inferior portion of the AVC in the same wt (G‐I) and Ztmem2 mutant (J‐L) hearts. V and A indicate the ventricle and atrium, respectively. For wt, n = 8; for Ztmem2, n = 6 PHENOTYPE:
|

ZFIN is incorporating published figure images and captions as part of an ongoing project. Figures from some publications have not yet been curated, or are not available for display because of copyright restrictions. |
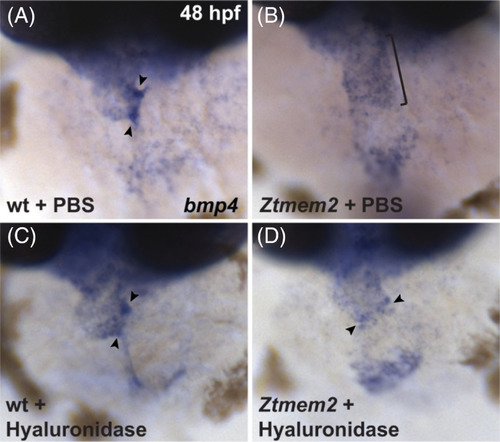
Hyaluronidase treatment can rescue the bmp4 expression pattern in Ztmem2 mutants. Expression of bmp4 in wt (A, C) and Ztmem2 mutant (B, D) embryos at 48 hpf, as in Figure 2. Pericardial injection of PBS did not alter the bmp4 expression pattern in wt (A) or Ztmem2 mutant (B) embryos (compare to Figure 2A,B). In contrast, pericardial injection of hyaluronidase restored the concentrated expression of bmp4 at the AVC in the majority of the injected Ztmem2 mutants (D, arrowheads; Table 3), but did not affect the bmp4 expression pattern in the majority of the injected wt sibling embryos (C, arrowheads; Table 3) |

Hyaluronidase treatment can restrict the distribution of Wnt signaling in Ztmem2 mutants. Three‐dimensional reconstructions depict immunofluorescent localization of Dm‐grasp (red) in the myocardium and GFP (green) driven by the Wnt signaling‐responsive reporter transgene Tg(7xTCF‐Xla.Siam:GFP)19 in wt (A, B, E, F) and Ztmem2 mutant (C, D, G, H) hearts at 48 hpf. In PBS‐injected wt embryos (A, B), Wnt signaling activity was restricted to the AVC (bracket in B). In PBS‐injected Ztmem2 mutants (C, D), the distribution of Wnt signaling activity extended beyond its normal boundaries (bracket in D). Although the cytoplasmic localization of the GFP reporter did not facilitate precise quantification, we conservatively estimate that Ztmem2 mutants typically exhibited ~50% more transgene‐expressing cardiomyocytes than did their wt siblings. Hyaluronidase treatment restricted the distribution of Wnt signaling activity in the majority of Ztmem2 mutants (G, H; Table 4), but did not affect the Wnt signaling distribution in most wt siblings (E, F; Table 4). We note that hyaluronidase treatment results in a particularly restricted pattern of Wnt signaling in Ztmem2 mutants (G, H), more restrained than that observed in wt (E, F). While we do not yet understand the basis for this restraint, we speculate that Tmem2 may be required for additional steps of AVC differentiation beyond the restriction of Wnt signaling, consistent with the prior observation that Ztmem2 mutants do not express the AVC differentiation marker spp1.8 In this scenario, while hyaluronidase treatment can rescue the restriction of Wnt reporter activity, it may not be sufficient to rescue additional roles of Tmem2 that are necessary to maintain robust levels of Wnt signaling within the AVC |
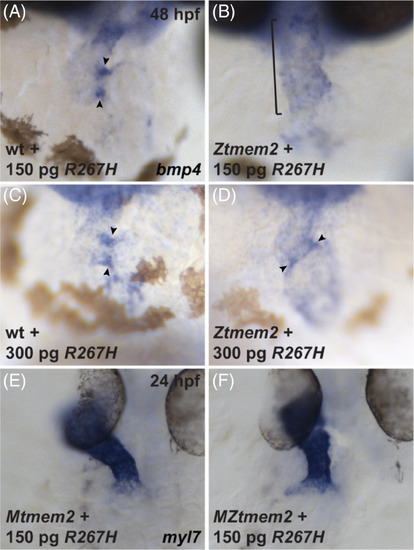
The R267H variant of Tmem2 exhibits hypomorphic activity in the context of AVC differentiation. Expression of bmp4 in wt (A, C) and Ztmem2 mutant (B, D) embryos at 48 hpf, as in Figure 2, and expression of myl7 in Mtmem2 (E) and MZtmem2 (F) mutant embryos at 24 hpf, as in Figure 4. Injection of 150 pg of R267H mRNA does not significantly alter the bmp4 expression pattern in Ztmem2 mutant embryos (B, bracket; Table 1), whereas doubling the dosage of R267H mRNA to 300 pg can rescue the bmp4 expression pattern in a majority of Ztmem2 mutants (D, arrowheads; Table 5). Injection of either dose of R267H mRNA does not affect the bmp4 expression pattern in wt siblings (A, C, arrowheads; Tables 1 and 3). In contrast to its limited activity in the context of AVC differentiation, injection of 150 pg of R267H mRNA efficiently rescues the cardiac fusion defects in MZtmem2 mutants (F; Table 2). Expression of R267H does not alter cardiac fusion in Mtmem2 siblings (E; Table 2) |