
- Title
-
Phenotypic Landscape of Schizophrenia-Associated Genes Defines Candidates and Their Shared Functions
- Authors
- Thyme, S.B., Pieper, L.M., Li, E.H., Pandey, S., Wang, Y., Morris, N.S., Sha, C., Choi, J.W., Herrera, K.J., Soucy, E.R., Zimmerman, S., Randlett, O., Greenwood, J., McCarroll, S.A., Schier, A.F.
- Source
- Full text @ Cell

Generation and Analysis of 132 Mutants for Schizophrenia-Associated Genes (A) Mutants were generated via Cas9 mutagenesis for genes found within and neighboring genomic loci linked to schizophrenia through genome-wide association. Manhattan plot image was adapted from Schizophrenia Working Group of the Psychiatric Genomics Consortium (2014). The resulting mutants were assessed for changes to brain morphology, brain activity, and behavior. Multiple zebrafishorthologs existed for 33 of the 132 genes (165 individual zebrafish genes), and both copies were mutated and assessed together. (B) Ricopili plot (https://data.broadinstitute.org/mpg/ricopili/) for multi-gene locus #3 of 108 from which five candidates (purple boxes) were selected for mutagenesis. (C) Ricopili plot for a gene (znf536) considered unambiguous because there are no other genes within 0.5 megabase on each side of the associated region, and the gene within the association region is brain-expressed. (D) Mutants made from 79 of 108 associated genomic loci. The locus rank reflects the statistical strength of the genetic association, with 1 being the most significantly associated. A region of 0.5–2 MB around each locus was analyzed, and genes outside of the region of linkage disequilibrium were selected for 19 of the 79 loci. Unambiguously associated genes are implicated strongly by previous literature, such as genes involved in glutamatergic neurotransmission, or are the only genes within or neighboring their locus (Table S1). (E) Mutations generated from Cas9 cleavage. Some mutant alleles consisted of several lesions if multiple gRNAs cleaved the genome independently, and all are included here. A range of mutations was recovered, tending to be either small (<15 bases) when a single gRNA cleaved or large (>100 bases) when a deletion spanned target sites of multiple gRNAs. (F) Protein sequence predicted to remain in mutants, based on sequence alignment identity. This analysis included both orthologs if the gene was duplicated (33 genes), for a total of 165 individual genes (163 in graph, as it does not include mir137 and one gene with unclear wild-type protein sequence length). The four mutants with >75% of the protein remaining did not have frameshifting mutations but did have phenotypes (Table S2), indicating that the protein function was disrupted. (G) Phenotypes in all 132 mutants based on analysis of brain activity signals and 71 behavioral assessments. See also Figure S1 and Figure S2for the cutoffs for classifying which mutants have phenotypes. (H) Phenotypes in mutants for the 29 unambiguously associated genes (Table S1). (I) Phenotype dimensions affected in mutants for 132 genes from 79 schizophrenia-associated loci. Quantification of brain activity, brain structure, and behavioral differences for mutants designated as having a phenotype (Figure S1, Figure S2, Figure 1G, Figure 3B, Table S2) was scaled for comparison between the three measures, with the weakest phenotype designated as 0 and strongest as 1. Measures below the cutoff for phenotype designation (Figure S1, Figure S2) are displayed in black. |

ZFIN is incorporating published figure images and captions as part of an ongoing project. Figures from some publications have not yet been curated, or are not available for display because of copyright restrictions. PHENOTYPE:
|

Whole-Brain Morphology Phenotypes in Zebrafish Mutants (A) Location of several major regions in the zebrafish brain (Randlett et al., 2015). (B) Comparison between brain activity and morphology data for all mutants. The rora mutant (orange triangle) represents the smallest (5.95) structural change designated as a phenotype, shown in panel C. Small signals that were not symmetrical were considered noise, compared to small but symmetrical changes as observed in the rora mutant. (C) Examples of structural differences in mutants calculated using deformation-based morphometry and displayed as sum-of-slices projections (Z- and X- axes). Brain images represent the significant differences in signal between two groups (Randlett et al., 2015), most often homozygous mutants versus heterozygous and/or wild-type siblings (Table S2). The number of animals used in all imaging experiments is available in Table S2 and on stackjoint.com/zbrain(website naming conventions for datasets described in STAR Methods). Brain maps were averaged from two independent clutches of larvae if the experiment was repeated and irreproducible signals were eliminated (STAR Methods). Raw imaging data examples (maximum projections) are shown for the foxg1 mutants, demonstrating that the forebrain in homozygous (HOM) mutants is underdeveloped. A reduction in forebrain size of heterozygous (HET) foxg1 mutants when compared to wild-type (WT) siblings can be quantified with deformation-based morphometry (sum-of-slices projection), although it is not readily apparent in the raw projection. (D) Fluorescent RNA in situ images for four mutants with altered brain morphology. |
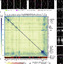
Whole-Brain Activity Phenotypes in Zebrafish Mutants The number of animals used in all imaging experiments is available in Table S2 (average N is 20 for mutant group, 28 for control group) and on stackjoint.com/zbrain (website naming conventions in STAR Methods). Brain maps were averaged from two independent clutches of larvae if the experiment was repeated and irreproducible signals were eliminated (STAR Methods). The white numbers in the upper right corner of images connect pERK activity maps, both of single genes (panels A and B, and Figure S5) and gene averages (panel E), to the heatmaps and to each other. See also Figure S4. See also Figure S5. (A) Brain activity phenotypes for genes that have been strongly implicated in schizophrenia by previous studies (Table S1). Sum-of-slices projections of significant differences between mutant and control groups of zebrafish larvae (Randlett et al., 2015). The cacna1c mutant phenotype shown is for heterozygous larvae because the homozygous mutant is embryonic lethal (Stainier et al., 1996). (B) Brain activity phenotypes for four genes that have been minimally studied and have unknown functions. (C) Percent overlap between mutant brain activity phenotypes was calculated between each image by comparing each brain activity signal to signal in the same location in all other mutant images. These overlaps were then sorted with hierarchical clustering using average linkage. Genes grouped together by this clustering are labeled. The direction of the change in brain activity was disregarded to maximize identification of affected brain regions, and because the direction of the genetic perturbation in human patients is not clear for most genes. The numbers of image stacks that were compared to calculate significant differences in brain activity for each mutant are available in Table S2. (D) Contribution of each of the four major brain divisions to the overall brain activity phenotype. The signal in each region was divided by the whole brain signal. Prior to dividing the signal in each region by the total signal, the regions were scaled relative to each other based on their respective sizes (rhombencephalon = ∗1, diencephalon = ∗1.76, mesencephalon = ∗1.42, telencephalon = ∗4.36). The original whole brain signal was separately scaled across all mutants with a phenotype, indicating the relevance of the signal in each of the regions. Measures below the cutoff for phenotype designation (Figure S1, Figure S2) are displayed in black. (E). Average signal for mutants with similar brain activity maps. Examples of individual maps that are included in these averages are labeled in panels A and B, as well as in Figure S5. The blue arrow highlights retinal arborization field AF7. (F) Two mutants with brain activity signals that overlap with both the forebrain (group 2) and the tectum (group 4). |

Nominating Candidates in Multi-Gene Loci by Phenotype The number of animals used in all imaging experiments is available in Table S2 and on stackjoint.com/zbrain. (A) Brain activity data (sum-of-slices projection) for lrrn3 and immp2lmutants. (B) Movement frequency for csmd3 mutants. P value = 0.0001; N = 30 +/- , 29 -/-. (C) Mutants with tectum (green arrow) and retinal arborization field AF7 (blue arrow, Mesencephalon___Retinal_Arborization_Field_7_AF7 on stackjoint.com/zbrain) phenotypes (representative slices). Mutants with retinal arborization field AF7 signal also display signal changes in the same direction in a small subregion of hypothalamus (orange arrow, Diencephalon___Hypothalamus_Gad1b_Cluster_3_Sparse located within cyan Diencephalon___Intermediate_Hypothalamus; see regions on stackjoint.com/zbrain). See also kmt2e, znf804a, cacnb2b, and ambra1(both areas decreased), and snap91, akt3b, and satb1 (both areas increased) on stackjoint.com/zbrain. (D) Prepulse inhibition phenotypes for five mutants. These mutant phenotypes are specific to the strong prepulse tap (Figure 2J) and do not represent a general increase in tap sensitivity to strong taps (right heatmap). Response features were calculated only on strong tap responses where the weak tap did not elicit movement. See also Figure S3. (E) Habituation phenotype of astn1 mutants. Response frequency to tap events occurring every two seconds is shown in left graph, and the magnitude of responses occurring during the habituation paradigm in the right graph. P value for frequency metric = 0.0018; merged p value for the day tap habituation 2 section = 0.015 with 20/47 significant metrics; N = 36 +/- , 18 -/-. |

Neurobiological Roles of Top Candidates (A) Example images of larvae after consuming fluorescently labeled paramecia. (B) Quantification of feeding behavior in tcf4 mutants by measurement of paramecia consumed. (C) T-distributed Stochastic Neighbor Embedding (t-SNE) (Hinton and Maaten, 2008) visualization of wild-type single-cell clusters obtained by clustering of 6 dpf forebrain cells. Clusters with substantial differences in znf536 mutants are highlighted in orange, purple, and blue. Cluster counts in mutant and wild-type are expressed as percent of the total cell number for each sample. See also Figure S6. (D) Dotplot (confusion matrix) showing the proportion of cells in the znf536 mutant forebrain that were classified to wild-type cluster labels. Each mutant forebrain type was assigned to a wild-type cluster label if > 13% of the trees in the Random Forest (RF) model (Breiman, 2001, Pandey et al., 2018) contributed to majority vote. |

Defining Imaging Phenotypes, Related to Figure 1, Figure 3, Figure 4 (A) Determination of cutoff for brain activity signals. Images are sum-of-slices intensity projections (Z- and X- axis). This cutoff for the log10(sum of pixels) was made so that mutants with small, specific, repeatable signals were classified as having phenotypes (gria1,forebrain). Full stacks of all individual repeats are available on stackjoint.com/zbrain. Many mutants with diffuse and sparse signals (likely noise) fell below the cutoff. Exceptions included klc1 and nrgn(shown here), which both had signals that reproduced in independent experiments (stackjoint.com/zbrain). (B) Examples of individual homozygous mutant and heterozygous control brains stained with phospho-Erk (clcn3, csmd1) or total-Erk (znf536) (maximum intensity projections). Twelve brains of each genotype are shown for the three genes. The localized reduction in forebrain activity observed in the clcn3 map is readily observable in the original images. In contrast, the widespread increased activity in the csmd1 map is not as discernable by eye, but is still noticeable in some areas (mesencephalon, rhombencephalon). The MapMAPPING protocol (Randlett et al., 2015) quantifies these less obvious differences. Significant voxels are defined using the Mann-Whitney U statistic Z score, intrinsically selecting for repeatable signals. Variability in brain activity between individuals is most likely due to subtle differences in the experiences of the larvae during the last ten minutes before fixation (the integration time of phospho-Erk). Structural changes in znf536mutants are discernable for the smaller size of the forebrain pallium and symmetric reduction in the cerebellum. The other small signals are likely noise, as they are asymmetric (Figure 3B; znf536 is in the “small changes and noise” group). |

Comparison between in situ RNA Localization and Brain Activity, Related to Figure 4 Stacks of all RNA-FISH images are available on http://stackjoint.com/basic/ and are tagged as “Thyme, 2018.” Probe sequences are available on http://genepile.com/scz_gwas108. At least three brains were imaged for each probe to confirm that the presented pattern was consistent (data available upon request). Brain areadescriptions are based on the Z-Brain masks (stackjoint.com/zbrain). znf536: forebrain showed in situ and phosphorylated-Erk (pErk) brain activity signal. grin2aa: cerebellum showed in situ and activity signal. elfn1a: retinal arborization field AF7, tectum, and forebrain showed in situ and activity signal. foxg1: forebrain showed in situ and activity signal. gria1a: forebrain showed in situ and activity signal. cacna1c:cerebellum and forebrain showed in situ and activity signal. hcn1:hindbrain showed in situ and activity signal. egr1: forebrain showed in situ and activity signal. roraa: tectum and hindbrain showed in situ and activity signal. nrgn: forebrain and hypothalamic cells showed in situ and activity signal. snap91: activity signal correlated to in situ signal showing expression throughout the brain. shisa9b: forebrain showed in situ and activity signal. znf804a: activity signal correlates to in situ signal showing expression throughout the brain. lrrn3: activity signal correlated to in situ signal showing expression throughout the brain. csmd1: activity signal correlated to in situ signal showing expression throughout the brain. cnnm2b: strong in situ signal in forebrain and torus semicircularis did not correlate to activity signal in tectum, retinal arborization field AF7, and hindbrain. csmd3: strong in situ signal in retinal arborization field AF7, tectum, and hindbrain did not correlate to diffuse and minimal activity signal. tcf4: strong in situ signal in forebrain and midbrain did not correlate to strong activity signal in tectum. luzp2:Strong in situ signal in tectum and cerebellum did not correlate to activity signal in hindbrain. tle3a: strong in situ signal in forebrain and tectum did not correlate to strong activity signal primarily in hindbrain. clcn3: strong in situ signal throughout the brain did not correlate to activity signal mainly in forebrain. grin2ab: strong in situsignal throughout the brain did not correlate to activity signal primarily in the tectum, retinal arborization field af7, and hindbrain. bcl11b:strong in situ signal throughout the brain did not correlate to activity signal in forebrain and cerebellum. |

Comparison of Brain Activity and Behavioral Datasets, Related to Figure 2 and Figure 4 (A) Comparison of baseline behavioral data across multiple time windows (Figure 2) with brain activity changes in the four major regions. Stimulus-driven behaviors (prepulse, dark flash response, etc) are not comparable to the baseline phospho-Erk data. There are several caveats to consider when comparing these two datasets. The behavioral data was collected on larvae in 96-well plates, isolated from interaction with any other larvae and constrained in their movement, while phospho-Erk staining was completed on a group of larvae in a large (150 mm) Petri dish. It is likely that the differences in these conditions would result in different brain states. The phospho-Erk dataset also represents only a single time point, the afternoon of 6 dpf, while the behavioral data was collected across two nights and days. Both the phospho-Erk and behavioral analyses only explore a small and relatively non-overlapping part of the possible behavioral landscape, and a more appropriate assessment would be to compare these two types of data collected under the same conditions for specific behaviors of interest. Further, finding similar differences in brain activity in a region does not necessarily mean that the exact same neurons are affected. (B) To search for connections between brain activity and behavior in an unbiased and quantitative way, we developed a procedure to compare the merged p values from baseline behavioral assays with the region-specific brain activity signal. Two relationships remained convincing after qualitative assessment (open circles) of the results: a relationship between reduced frequency of motion and a reduced activity in the telencephalon (C), and a relationship between increased activity in the rhombencephalon and increased preference for the well edge (D). Briefly, we binarized both the brain activity and behavioral data and conducted Fisher exact enrichment analyses, followed by combining the resulting p values for all time windows to look for relationships that were consistent across time and therefore more likely real. For the brain activity data, we focused on the 61 largest regions based on their size in pixels, and normalized the total number of pixels with signal by dividing by the region size. Regions with less than 1% activity were designated as 0 and regions with greater than 1% signal were designated as 1. This binarization was not optimal and certainly resulted in false positive designations of signal (filled-in circles represent contributions to quantitative significance that were potentially false designations, compared to open circles where binarization more accurately represented the data). For the behavioral data, any assay with a significant merged p value (< 0.05) was designated as 1. Following initial enrichment analyses across a matrix of the 61 regions and 45 baseline time windows, the resulting Fisher p values were combined across the time windows using Fisher’s method. The most significant combined p values are shown for the comparisons of both increased behavior and decreased with increased and decreased binarized brain activity signal. If a comparison is not shown, no combined p values of substantial significance were identified. (C) Brain activity maps for the forebrains of eight mutants that have both convincing telencephalon activity reductions and a consistent reduction in baseline frequency of motion (open circles, A). (D) Three examples of mutants with an increased preference for the well edge. Correspondingly, we observed substantial increases in brain activity in these mutants, specifically in the hindbrain region for elfn1(see also Figure S5 brain activity map for only elfn1a) and hcn1. Additional examples are highlighted with open circles (A) and are available to view as stacks on stackjoint.com/zbrain. Intriguingly, several of these mutants have also been implicated in epilepsy (references in Table S1), another neuropsychiatric disease that may share genetic underpinnings with schizophrenia. |
Reprinted from Cell, 177(2), Thyme, S.B., Pieper, L.M., Li, E.H., Pandey, S., Wang, Y., Morris, N.S., Sha, C., Choi, J.W., Herrera, K.J., Soucy, E.R., Zimmerman, S., Randlett, O., Greenwood, J., McCarroll, S.A., Schier, A.F., Phenotypic Landscape of Schizophrenia-Associated Genes Defines Candidates and Their Shared Functions, 478-491.e20, Copyright (2019) with permission from Elsevier. Full text @ Cell