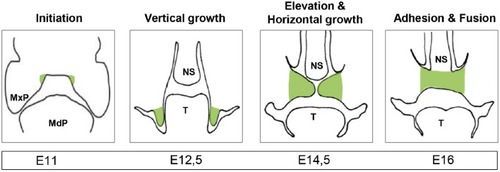
Palatogenesis. Illustration of the developing secondary palate (highlighted in green) through frontal sections of a mouse embryo with the timing of the relevant stages below. MxP, maxillary process; MdP, mandibular process; NS, nasal septum; T, tongue; highlighted in green: palatal shelves.
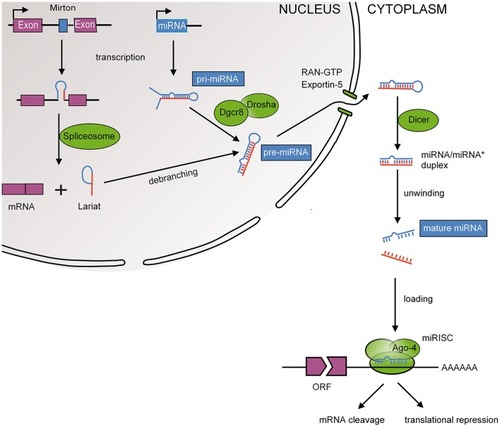
MiRNA biogenesis. Mature miRNAs are encoded in the genome and form after a series of enzymatic cleavages from two possible precursor molecules; primary miRNAs (pri-miRNAs) or Mirtrons. Pri-miRNAs, following the canonical pathway, are transcribed as long hairpin RNAs that are recognized by the RNA-binding DiGeorge syndrome critical region 8 protein (DGCR8). Many pri-miRNAs are often transcribed simultaneously due to clustering of several miRNA genes (Ambros et al., 2003). DGCR8 then directs the RNase III endonuclease DROSHA to cleave the base of the hairpin to produce ~70 nucleotide hairpins known as pre-miRNA. Mirtrons, following the non-canonical pathway, bypass the microprocessor as they are transcribed as part of the introns of protein coding genes and are as such spliced by the spliceosome (Berezikov et al., 2007). Splicing also produces ~70 nucleotide hairpins known as pre-miRNA. The pre-miRNA is transported to the cytoplasm by exportin 5 where it is cleaved by another RNase III endonuclease known as DICER to ~20 nucleotide miRNA duplexes with protruding 2 nucleotide 3′ ends. The resulting mature miRNA is released and a guiding strand is incorporated into the RNA-induced silencing complex (RISC).
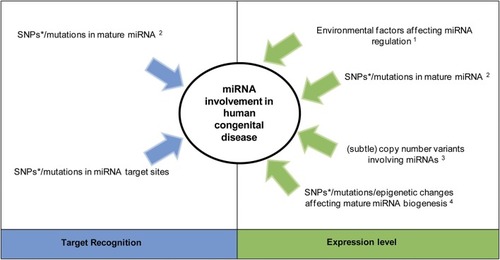
MiRNA involvement in congenital disease. Alterations affecting miRNA activity by changing target recognition or modulating their expression. *Single nucleotide polymorphisms (SNPs) within miRNAs are likely involved in complex disease (common disease, common variant hypothesis). 1Environmental factors can directly or indirectly regulate miRNA expression independent of any germline miRNA alteration (Zhao et al., 2008). 2A SNP/mutation within the miRNA seed sequence can alter both its processing and target recognition, while a change outside the seed sequence only alters miRNA processing (Duan et al., 2007). 3 Large copy number variants lead to syndromes while subtle ones (those only detectable via molecular methods) are predicted to be involved in complex diseases (Shelling and Ferguson, 2007). 4Germline alterations of regulators belonging to one of the two miRNA biogenesis/ processing pathways (i.e., the pathways involving mature miRNA generation from pri-miRNAs or Mirtrons, see Figure 2) will only change the expression level of mature miRNAs being generated through this pathway (Finnegan and Pasquinelli, 2013). Epigenetic changes in this context refer to functional changes without a change in the DNA sequence, such as methylation and histone modification.
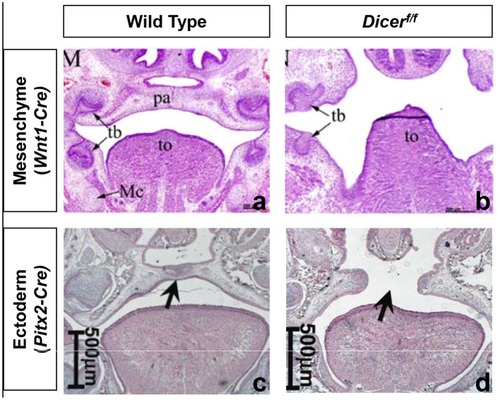
Homozygous conditional deletion of Dicer in neural crest derived mesenchyme and oral ectoderm. Coronal sections of E18.5 wild-type and Wnt1-Cre; Dicerf/f(a,b) and E16.5 wild type and Pitx2-Cre; Dicerf/f(c,d). Black arrow: (left) palate. (a,b) Adapted from Nie et al. (2011). (c,d) Adapted from Cao et al. (2010). pa, palate; tb, tooth bud; Mc, Meckel's cartilage; To, tongue.
Acknowledgments
This image is the copyrighted work of the attributed author or publisher, and
ZFIN has permission only to display this image to its users.
Additional permissions should be obtained from the applicable author or publisher of the image.
Full text @ Front. Physiol.
Your Input Welcome
Thank you for submitting comments. Your input has been emailed to ZFIN curators who may contact you if
additional information is required.
Oops. Something went wrong. Please try again later.