
- Title
-
Mutations disrupting the ordering and topographic mapping of axons in the retinotectal projection of the zebrafish, Danio rerio
- Authors
- Trowe, T., Klostermann, S., Baier, H., Granato, M., Crawford, A.D., Grunewald, B., Hoffman, H., Karlstrom, R.O., Meyer, S.U., Muller, B., Richter, S., Nüsslein-Volhard, C., and Bonhoeffer, F.
- Source
- Full text @ Development

Mutations affecting the sorting of axons into the two branches of the optic tract. (A) In wild-type animals, nasodorsal axons (green) grow in the ventral branch of the optic tract (arrow). (B) In box mutant fish, nasodorsal axons grow in the ventral (arrow) and in the dorsal branch (arrowhead) of the optic tract. (C) Another box mutant fish. Nasodorsal axons that enter the tectum through the dorsal branch continue their dorsal trajectory on the tectal lobe (arrow). Near the posterior margin of the lobe, they turn and terminate retinotopically, posteroventrally (*). Some of the ectopic nasodorsal axons leave the contralateral tectal lobe and grow to the ipsilateral one (arrowhead). (D) Trajectory of middorsal RGC axons of a box mutant. Dorsal view of the contralateral tectal lobe. Middorsal axons grow in both branches of the optic tract and on the dorsal and the ventral side of the tectal lobe (arrowheads). Ectopic middorsal axons make a sharp turn middorsally (arrow) and grow to their retinotopic midventral target area (*). (E) Nasodorsal axons in a dak mutant. As in box, these axons grow in both branches of the optic tract. Ectopic nasodorsal axons grow around the dorsal side of the tectal lobe (arrowhead), but terminate retinotopically, posteroventrally (*). The nasodorsal axons on the dorsal side of the lobe are much more strongly fasciculated than the ones on the ventral side (arrows). (F) Confocal image of the retinotectal projection of a pic mutant. Nasodorsal axons grow in both branches of the optic tract (arrows). The nasodorsal axons in the dorsal branch and some temporoventral axons turn and grow to the ipsilateral tectal lobe (arrowhead). Here, the temporoventral axons terminate retinotopically (x), while the nasodorsal axons first grow around the dorsal margin of the lobe and then leave it to grow towards the contralateral lobe again (double arrowhead). Scale bars, 0.1 mm. PHENOTYPE:
|

Transverse sections through the tectal lobes of a wild-type fish (A) and three mutants (B,C,D). Before sectioning, nasodorsal RGCs were labeled with DiI and the dye was photoconverted. Medial is to the left, dorsal is up. (A) A wild-type fish. The termination field of the nasodorsal axons occupies the ventral half of the tectum. Black arrows mark the ventral margin of the termination fields in all four pictures, black arrowheads the dorsal margin. The dorsal margin of the tectal lobes is labeled by white arrowheads, the ventral margin by white arrows. The black bar marks the superficialdeep axis of the tectum. (B) In box mutants, the termination field of the nasodorsal axons occupies the ventral half of the tecum, as in wild-type fish. The tectum is compressed along the superficial-deep axis. (C) In nev fish, the termination field of nasodorsal axons reaches into the dorsal side of the tecum. It is compressed along its superficial-deep axis. (D) In woe mutants, nasodorsal axons terminate in two separate fields posteroventrally and posterodorsally in the tectum. The dorsal field is oriented more towards the center of the neuropil than towards the dorsal margin. Scale bars, 50 µm. |

Mutations affecting the mapping of RGC axons. All images shown in this figure are projections of optical sections obtained with a confocal microscope. (A) Retinotectal projection of a wild-type fish. The absolute position of the termination areas on the tectum cannot be seen, but the temporoventral axons (red) terminate anterior and dorsal of the nasodorsal axons (green), as is typical for a wild-type projection. For all the confocal images shown, the absolute positions of the termination fields has been ascertained using conventional fluorescence microscopy. (B) Retinotectal projection of another wild-type fish. In this animal, red axons are of nasoventral origin and terminate in the posterodorsal tectum (x). Note that the dorsal and ventral axons, although both originating in the nasal retina, still occupy different parts of the optic nerve (arrow). (C) In nev mutants, nasodorsal axons terminate posteroventrally (*) and posterodorsally (*′) in the tectum. Temporoventral axons terminate retinotopically in the anterodorsal tectum (x). The nasodorsal axons grow through both the ventral (arrow) and the dorsal (arrowheads) branch of the optic tract. RGC axons are already missorted in the optic nerve. Some of the temporoventral axons grow ectopically in the anterior portion of the optic nerve (double arrowheads). The inlet shows the optic nerve of another nev mutant individual showing a similar phenotype. (D) Another nev mutant fish. This picture shows more clearly that the nasodorsal axons grow in the ventral (arrow) and the dorsal (arrowhead) branches of the optic tract. The nasodorsal axons terminate again in the posteroventral (*) and posterodorsal (*′) tectum. Nasoventral axons (red) terminate retinotopically in the posteroventral tectum (x). (E) Nasoventral axons (red) in nev mutants always terminate correctly in the dorsal tectum (x), but reach their target area on meandering paths (arrowheads). A subpopulation of the nasoventral axons grows ectopically in the posterior portion of the optic nerve (arrow). Compare with B. In this individual, the nasodorsal axons terminate correctly in the ventral tectum, but the termination area is larger and fuzzier than in wild-type fish. (F) A woe mutant fish. Nasodorsal axons terminate on the ventral (*) and the dorsal (*′) side of the tectum. Note that the majority of the nasodorsal axons appear to terminate at the ectopic dorsal side. Temporoventral axons terminate retinotopically in the anterodorsal tectum (x). The RGC axons are normally sorted in the optic nerve (arrow) and tract (arrowhead), but the nasodorsal and the temporoventral fascicles cross each other further away from the eye than usual. The two branches of the tract itself are less widely searated than in wild-type animals. G: Another woe mutant fish. Nasodorsal axons (green) terminate again in the posteroventral (*) and the posterodorsal (*′) tectum. Nasodorsal axons split only upon reaching the tectum (arrow). Ectopic nasodorsal axons grow in several fascicles along the dorsal margin of the tectum (arrowheads). (H) Retinotectal projection of a gna mutant fish. The termination area of the nasodorsal axons is enlarged and encompasses posteroventral (*) and anteroventral (*′) parts of the tectum. The nasodorsal fascicle splits abruptly into several smaller fascicles upon reaching the tectum (arrows), some of which turn sharply dorsally before continuing to the posterior tectum (arrowhead). Temporoventral axons terminate correctly in the anterodorsal tectum (x). The sorting of the axons along the optic nerve and tract is normal. (I) In some gna mutant fish, nasodorsal axons terminate not only anteroventral (*′) and posteroventral (*), but also posterodorsal (*′′). (J) A mao mutant fish. The termination area, especially of the nasodorsal axons (*), is larger and fuzzier than in wild type. m, melanophore. Scale bar, 0.1 mm. |
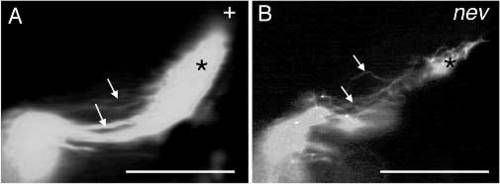
Nasoventral axons of a wild-type fish and of a nev mutant. Dorsal views of the tectal lobes, anterior is to the left. (A) In wildtype fish, nasoventral axons grow in straight, smoothly curved trajectories (arrows) on the dorsal side of the tectal lobe and terminate posterodorsally. (B) A nev mutant fish. Nasoventral axons terminate retinotopically (*), but they grow on tortous paths (arrows). m, melanophore. Scale bars, 0.1 mm. PHENOTYPE:
|

Mutations affecting tectal size. (A) Retinotectal projection of a wild-type fish. (B) Retinotectal projection of a noi mutant. The tectum (dotted line) is smaller than in wild-type fish. Temporoventral axons (arrow) project to the posterior margin of the tectal remnant (arrowheads). (C) Nasodorsal axons of a noi mutant. They pass the posterior end of the tectal remnant (dotted line) and grow caudally (arrows). (D) Retinotectal projection of a tin mutant. The tectum (dotted line) is tiny. Even in this tiny tectum, nasodorsal axons (green) project posterior and ventral of the temporoventral axons (red). m, melanophore. Scale bars, 0.1 mm. PHENOTYPE:
|

Mutations affecting the formation of termination fields. (A) Retinotectal projection of a wild-type fish. (B) Retinotectal projection of an esr mutant. Retinal axons form a thickening right behind the optic papilla (arrow). Although many axons are still seen in the optic tract, the termination fields of the RGC axons on the tectum are tiny (arrowheads). Nasodorsal axons are not seen in the posterior tectum (*). (C) Nasodorsal axons of esr. In this case, the nasodorsal axons reach the posterior margin of the tectum (approximate position marked by arrowheads). Their termination field is spread out and covers a larger area than usual. (D) Nasodorsal axons of blu. Upon reaching their target area in the posteroventral tectum, they arborize in a cauliflower-like fashion (arrowheads). (E) Retinotectal projection in a brd mutant. Many nasodorsal axons terminate prematurely already in the midventral tectum (arrowhead). Nasodorsal axons in the posteroventral tectum are marked by an arrow. m, melanophore. Scale bars, 0.1 mm. PHENOTYPE:
|
