
Fig. S2
|
Fig. S2
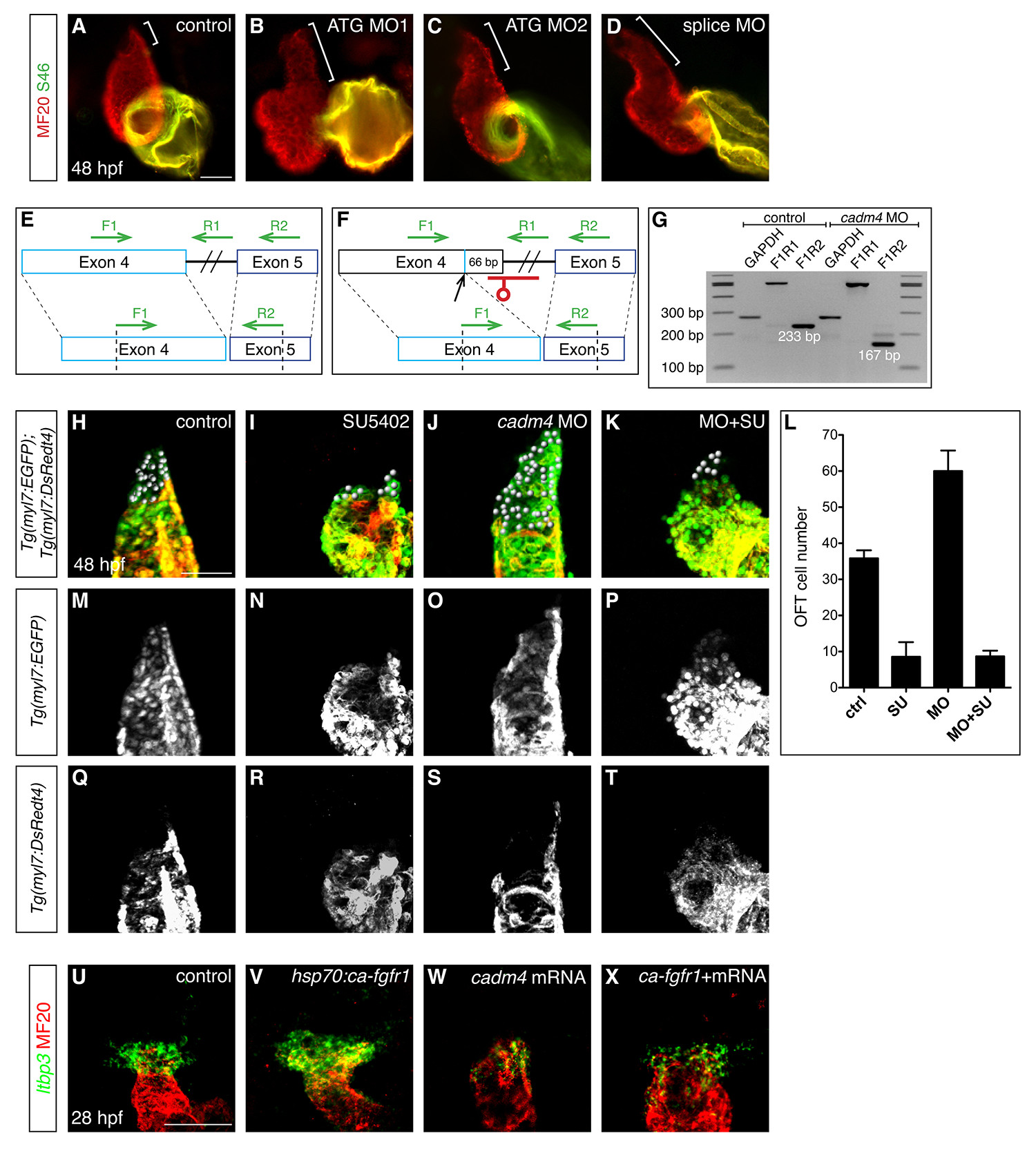
Multiple anti-cadm4 MOs cause OFT elongation, and cadm4 plays a potent role in limiting the formation of Fgf-dependent OFT cells.
(A-D) Hearts at 48 hpf, as in Fig. 1A. Compared to the OFT length in control embryos (bracket, A), embryos injected with any of three anti-cadm4 MOs exhibit OFT elongation (brackets, B-D).
(E-G) Injection of a splice-blocking MO efficiently interferes with cadm4 splicing. Schematics (E,F) depict a portion of cadm4 pre-mRNA and mRNA. In contrast to normal splicing between exons 4 and 5 (E), embryos injected with the splice MO utilize a cryptic splice donor site, 66 bp upstream of the usual donor site (F). This alteration is predicted to remove 3 amino acids from the end of the second Ig-like domain and 19 amino acids from the region linking the second and third Ig-like domains. (G) The effectiveness of the splice MO is visualized by comparing RT-PCR amplification products using cDNA from control and splice MO-injected embryos. Primer pair F1R1 amplifies 517 bp of unspliced cadm4 pre-mRNA, and primer pair F1R2 amplifies 233 bp of normally spliced cadm4 mRNA or 167 bp of abnormally spliced cadm4 mRNA. An amplification product from the housekeeping gene gapdh is used as a control. (H-K, M-T) Partial reconstructions of Tg(myl7:EGFP);Tg(myl7:DsRedt4) hearts at 48 hpf, arterial pole up, distinguish early-differentiating and late-differentiating CMs, as in Fig. 2J-R. Spheres indicate late-differentiating CMs (green and not red) in (H) control embryos, (I) embryos treated with SU5402 from 24-48 hpf, (J) cadm4 morphants, and
(K) cadm4 morphants treated with SU5402 from 24-48 hpf.
(L) Number of late-differentiating CMs present at 48 hpf in control embryos (n=6), embryos treated with SU5402 from 24-48 hpf (n=9), cadm4 morphants (n=8), and cadm4 morphants treated with SU5402 from 24-48 hpf (n=9). Treatment of cadm4 morphants with SU5402 reduces the formation of late-differentiating CMs to a number similar to that seen in uninjected embryos treated with SU5402. Therefore, formation of excess late-differentiating OFT cells in cadm4 morphants is dependent upon Fgf signaling.
(U-X) In situ hybridization depicts expression of ltbp3 (green) in progenitor cells adjacent to the differentiated myocardium (MF20, red); partial reconstructions of dorsal views at 28 hpf, arterial pole up. In comparison to control embryos (U), the ltbp3- expressing progenitor population is expanded in Tg(hsp70:ca-fgfr1) embryos (V) that experience increased Fgf signaling following heat shock at 22 hpf. In embryos overexpressing cadm4 (W), ltbp3 expression is reduced in a manner reminiscent of the effect of cadm4 overexpression on mef2cb expression (Fig. 3C). Expression of ltbp3 is similarly reduced in heat-shocked Tg(hsp70:ca-fgfr1) embryos that are also overexpressing cadm4 (X). Therefore, high levels of cadm4 activity can override the influence of elevated Fgf signaling on the OFT progenitor population.
Scale bars: 50 μm.