
- Title
-
Disrupting the Repeat Domain of Premelanosome Protein (PMEL) Produces Dysamyloidosis and Dystrophic Ocular Pigment Reflective of Pigmentary Glaucoma
- Authors
- Hodges, E.D., Chrystal, P.W., Footz, T., Doucette, L.P., Noel, N.C.L., Li, Z., Walter, M.A., Allison, W.T.
- Source
- Full text @ Int. J. Mol. Sci.
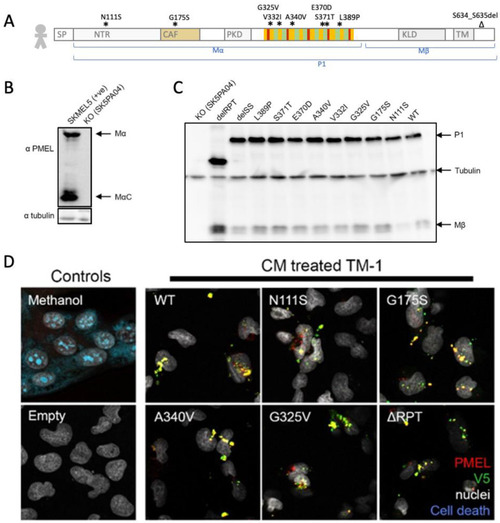
Human PMEL is engulfed by trabecular meshwork cells, providing a |
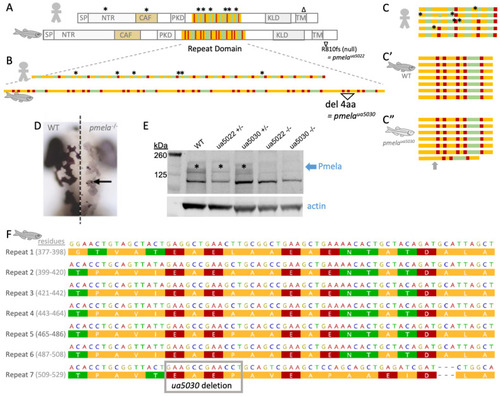
A zebrafish model of pigmentary glaucoma engineered via subtle mutation to the repeat region of the |
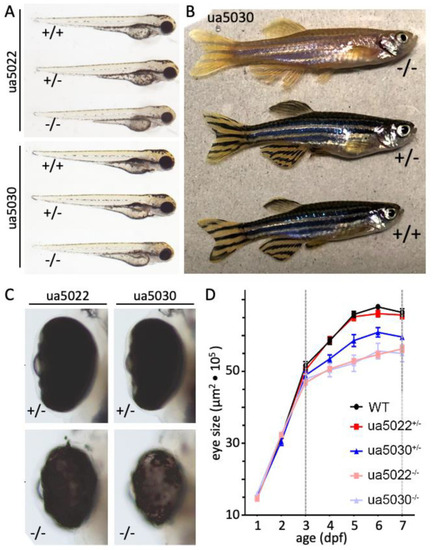
|
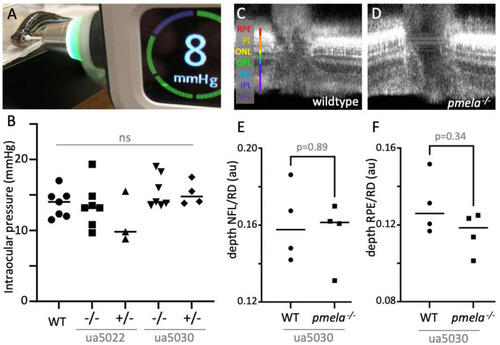
Adult |
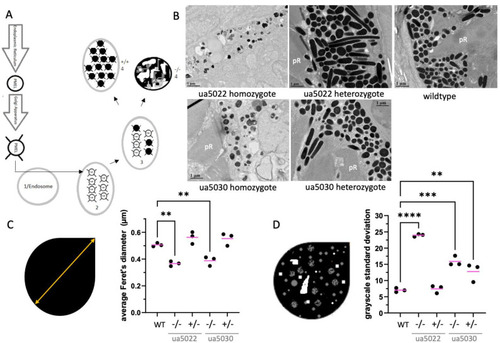
The repeat region of the Pmela protein is required for the elongation of melanosomes and the even distribution of melanin within melanosomes. ( |
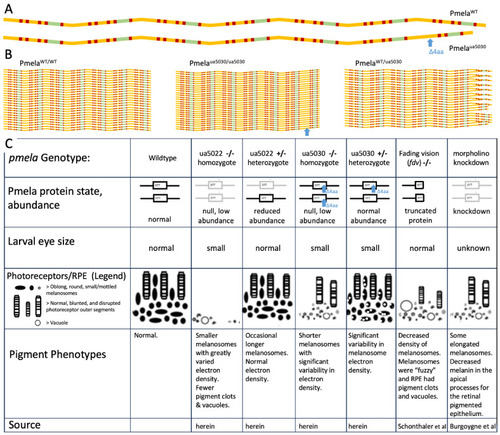
Summary of |