
- Title
-
CFTR Depletion Confers Hypersusceptibility to Mycobacterium fortuitum in a Zebrafish Model
- Authors
- Johansen, M.D., Kremer, L.
- Source
- Full text @ Front Cell Infect Microbiol

|
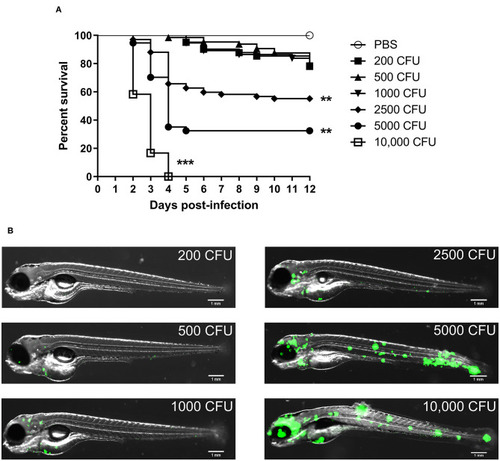
Zebrafish embryos are susceptible to |

Zebrafish infected with |

Lipoclodronate macrophage depletion results in lethal |

CFTR ablation leads to rapid larval death and uncontrolled bacterial expansion. |

A hypothetical schematic summarizing the pathogenesis of |