
- Title
-
Systematic optimization and prediction of cre recombinase for precise genome editing in mice
- Authors
- Erhardt, V., Hartig, E., Lorenzo, K., Megathlin, H.R., Tarchini, B., Hosur, V.
- Source
- Full text @ Genome Biol.

Optimizing Cre-Lox recombination: factors affecting efficiency and reliability. a The Cre-Lox system, consisting of the enzyme Cre and its specific DNA sequence lox, enables spatial-temporal control over gene expression in mice. It is widely used in research, allowing researchers to study the function of specific genes with precise control over expression time and location, deepening our understanding of biological processes. Cre-Lox technology has a notable limitation?the unpredictability of efficiency of Cre-mediated recombination and mosaicism (i.e., incomplete recombination?the targeted genetic changes do not occur uniformly across the targeted tissue). Although a high recombination efficiency is in most cases the desired outcome, for some biological questions, genetic mosaicism is of physiological relevance. Our lack of understanding of how various considerations affect recombination efficiency, especially when using tissue-specific Cre drivers, undermines the reliability of the Cre-Lox system, leading to variable experimental outcomes that may not align optimally with the investigator?s intention. b We systematically examined the role of various factors, including the Cre-driver strain, inter-loxP distance, Cre breeder age at the time of mating, zygosity of the floxed allele (heterozygous versus homozygous floxed allele), wildtype versus mutant loxP sites, and the various genetic loci of floxed alleles across the genome on Cre-recombination efficiency and mosaicism. |

Overview of the pipeline and the methods used to assess the efficiency of recombination and mosaicism. a In this study, we implemented a thorough characterization pipeline comprising several crucial steps. Our initial focus was on characterizing three unique global Cre-driver strains, achieved by mating female Cre-driver strains with male R26-floxed strains. The resulting F1 offspring were genotyped using either standard PCR or probe-based assays. The genetic material for these assays was derived from tail tips of E17.5 stage embryos or postnatal P1 to P3 pups. For tissue-specific analysis of recombination and mosaicism, we conducted histological and immunohistochemical analyses using whole embryos from E15.5 to E17.5 or P2 pups, obtained from the mating of female floxed and male tissue-specific Cre-strains. b The percentage of progeny recombination was determined by dividing the number of litters showing complete recombination by the total number of litters and multiplying it by 100. The percentages of progeny mosaicism and floxed patterns were determined in a similar fashion. |

Cre-driver strain impact on recombination and mosaicism at different inter-loxP distances. a Schematic representation of a 0.8-kb floxed allele and a post-cre recombined allele. b Male homozygous floxed mice were bred with female Ella-cre, CMV-cre, or Sox2-cre mice. Note that Ella-cre and CMV-cre are homozygous, and Sox2-cre is hemizygous for the Cre allele. The F1 offspring were then genotyped using PCR from tail DNA to identify floxed and recombined alleles. Offspring with complete recombination were categorized as recombined, those with no recombination as still floxed, and those with both recombined and floxed alleles as mosaic. ?n? denotes the number of offspring genotyped. c Schematic representation of an 8-kb floxed allele and a post-cre recombined allele. Note that there is a 10-fold difference in the inter-loxP distance compared to the floxed allele analyzed in (a). d Homozygous floxed male mice were mated with either female Ella-cre, CMV-cre, or Sox2-cre mice, and the F1 offspring were genotyped to screen for floxed and recombined alleles. e The lengths of all protein-coding genes in mice are arranged in ascending order. Gene lengths varied widely from 0.066 kb to 2270 kb. The median length of protein-coding genes in mice is 15.8 kb. f Homozygous floxed male mice with a 10 kb inter-loxP distance were mated with Sox2-cre mice, and the F1 offspring were genotyped to screen for floxed and recombined alleles. Only 22% of the offspring showed complete recombination, while 41% were mosaic, and the remaining 37% showed no recombination. g Homozygous floxed male mice with a 12 kb inter-loxP distance were mated with Sox2-cre mice, and the F1 offspring were genotyped. None of the offspring showed complete recombination; however, 14% of the offspring were mosaic. h Homozygous floxed male mice with a 15 kb inter-loxP distance were mated with Sox2-cre mice, and the F1 offspring were genotyped using PCR from tail DNA to screen for floxed and recombined alleles. All the offspring had intact floxed alleles and showed no signs of recombination or mosaicism. |

Impact of inter-loxP distances on Cre-mediated recombination and mosaicism in DIO mouse strains: from genotyping to single-cell analysis. a Study design for generating conditional alleles at a fixed locus (R26) using the Cas9+Bxb1 toolbox. The Cas9+Bxb1 toolbox allows for accurate and efficient integration of large DNA constructs into a specific location, achieving higher efficiency compared to Cas9-mediated homology-directed repair (HDR). Bxb1 uses attP and attB attachment sites for DNA transgenesis. In our study, we strategically placed an attP attachment site within the R26 safe harbor locus of the B6/J mouse strain using CRISPR/Cas9-mediated HDR. This approach enabled us to seamlessly integrate DNA constructs with DIO alleles ranging from approximately 1 to 8 kb in size. Consequently, we successfully generated conditional knockout mice with single-copy transgenic modifications. b, c, and d Homozygous floxed male mice were mated with either female Ella-cre (b), CMV-cre (c), or Sox2-cre (d) mice, and the F1 offspring were genotyped using PCR from tail DNA to screen for floxed and recombined alleles. Offspring with complete recombination were categorized as recombined, those with no recombination as still floxed, and those with both recombined and floxed alleles as mosaic. The x-axis denotes the inter-loxP distance, and the y-axis denotes the percentage of recombination or mosaicism. The number of offspring genotyped to assess the percent of three possible outcomes: complete recombination, mosaicism, or no recombination are summarized in Additional file 1: Fig. S3b. e Mosaicism at the single-cell level: Confocal image of an 8-kb Myo15a-Halo DIO floxed allele x CMV-cre F1 cochlear sensory epithelium at postnatal day 2. Phalloidin stains filamentous actin (F-Actin), labeling the sensory hair cell cytoskeleton. TMR-conjugated Halo Tag ligand labels Halo-Myo15a, the product of recombination. Halo-Myo15a enriches at the tips of hair cell cytoskeletal projections. White asterisks indicate recombined cells expressing Halo-Myo15a. Yellow arrowheads indicate cells that escaped recombination, where the allele is still floxed. f Quantification of percentages from four different pups as points and the averages by bar plot. |

Inter-loxP distance and site type influence on Cre-mediated recombination efficiency and mosaicism. a Male homozygous floxed mouse strains, with a fixed inter-loxP distance of 6.9 kb at different loci and carrying either wildtype loxP sites or mutant loxP sites (lox71 and lox66), were bred with female Ella-cre, CMV-cre, or Sox2-cre mice. b Male homozygous floxed mouse strains, with an inter-loxP distance of 2.7 kb and 2.9 kb at different loci, having wildtype loxP sites and mutant lox71/66 sites, respectively, were bred with female Sox2-cre mice. c Male homozygous floxed mouse strains, with an inter-loxP distance of 6.9 kb or 2.9 kb at the same locus and with mutant loxP sites (lox71 and lox66), were bred with female Sox2-cre mice. In Figures 5a, b, and c, F1 offspring were genotyped using PCR from tail DNA to detect floxed, mosaic, and recombined alleles. Offspring with both recombined and floxed alleles were considered mosaic. Note that percentages were rounded to the nearest whole number. d Rhbdf1-/- Rhbdf2-/- double KO mice exhibit the ?eyelids open at birth? phenotype (indicated by arrows) and post-natal lethality [43]. However, when we generated Rhbdf1 conditional KO mice with mutant loxP sites (lox71 and lox66) and an inter-loxP distance of 6.9 kb (Rhbdf1flox/flox-6.9kb) , and crossed with Sox2-cre mice, we were unable to generate Rhbdf1flox/flox-6.9kb Rhbdf2-/- mice carrying the recombined Rhbdf1 allele (top panel?mosaic). Conversely, reducing the inter-loxP distance of the Rhbdf1flox/flox-6.9kb conditional KO allele from 6.9 kb to 2.9 kb using CRISPR/Cas9-mediated deletion of up to 3 kb (Rhbdf1flox/flox-2.9kb), and crossing them with Rhbdf2-/- and Sox2-cre mice, resulted in the generation of the Rhbdf1 recombined allele. This led to in utero lethality in Rhbdf1flox2.9/flox2.9 Rhbdf2-/- Sox2-cre mice (bottom panel?complete recombination). e The ?eyelids open at birth? phenotype (indicated by arrows) was observed in the Rhbdf1flox/flox-2.9kb Rhbdf2-/- KRT14-cre mice (bottom panel?complete recombination) but not in the keratinocyte-specific knock-out Rhbdf1flox/flox-6.9kb Rhbdf2-/- KRT14-cre mice (top panel?mosaic). f In utero lethality was only observed with the Rhbdf2-/- Rhbdf1flox/flox-2.9kb conditional KO allele (bottom panel?complete recombination), not with the Rhbdf2-/- Rhbdf1flox/flox-6.9kb allele (top panel?Mosaic), when endothelial-specific Cdh5-cre was used. |
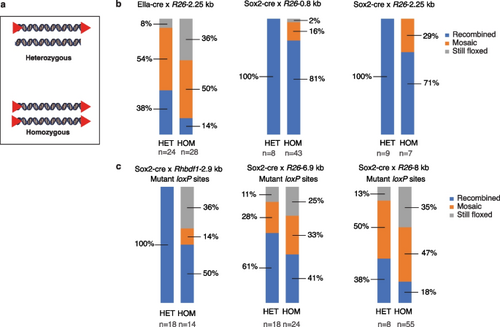
Impact of the zygosity of the floxed allele on recombination efficiency. a Schematic representation of the heterozygous (HET) versus homozygous (HOM) floxed alleles. b and c Male homozygous and heterozygous floxed mouse strains with either wildtype loxP (b) or mutant loxP (c)?R26-0.8 kb, R26-2.25 kb, Rhbdf1-2.9 kb, R26-6.9 kb, and R26-8 kb?were bred with female Ella-cre or Sox2-cre mice. The F1 offspring were then genotyped using PCR and categorized as recombined, still floxed, or mosaic. Note that the percentages were rounded to the nearest whole percent. |

Age-dependent recombination efficiency in mouse breeders and its impact on Cre-mediated recombination of floxed alleles. a Representation of mouse breeders ranging in age from 5 to 38 weeks was used to determine recombination efficiency. b Effects of age on Cre-mediated recombination of four different floxed alleles. Each data point represents the percentage of recombination in a litter of pups, coded by cross of origin, as labelled on plot. X-axis is the age (in weeks) of the Cre-driver breeder at the time of progeny birth. |

Extensive characterization of homozygous conditional knockout mouse strains using Sox2-cre recombination. Twelve different male homozygous conditional KO strains, with inter-loxP distances ranging from 0.8 to 3 kb at various loci across the mouse genome, were crossed with the Sox2-cre strain. The F1 offspring were then genotyped to identify floxed, mosaic, and recombined alleles. Note that the percentages were rounded to the nearest whole percent |