
|
Fig. 7
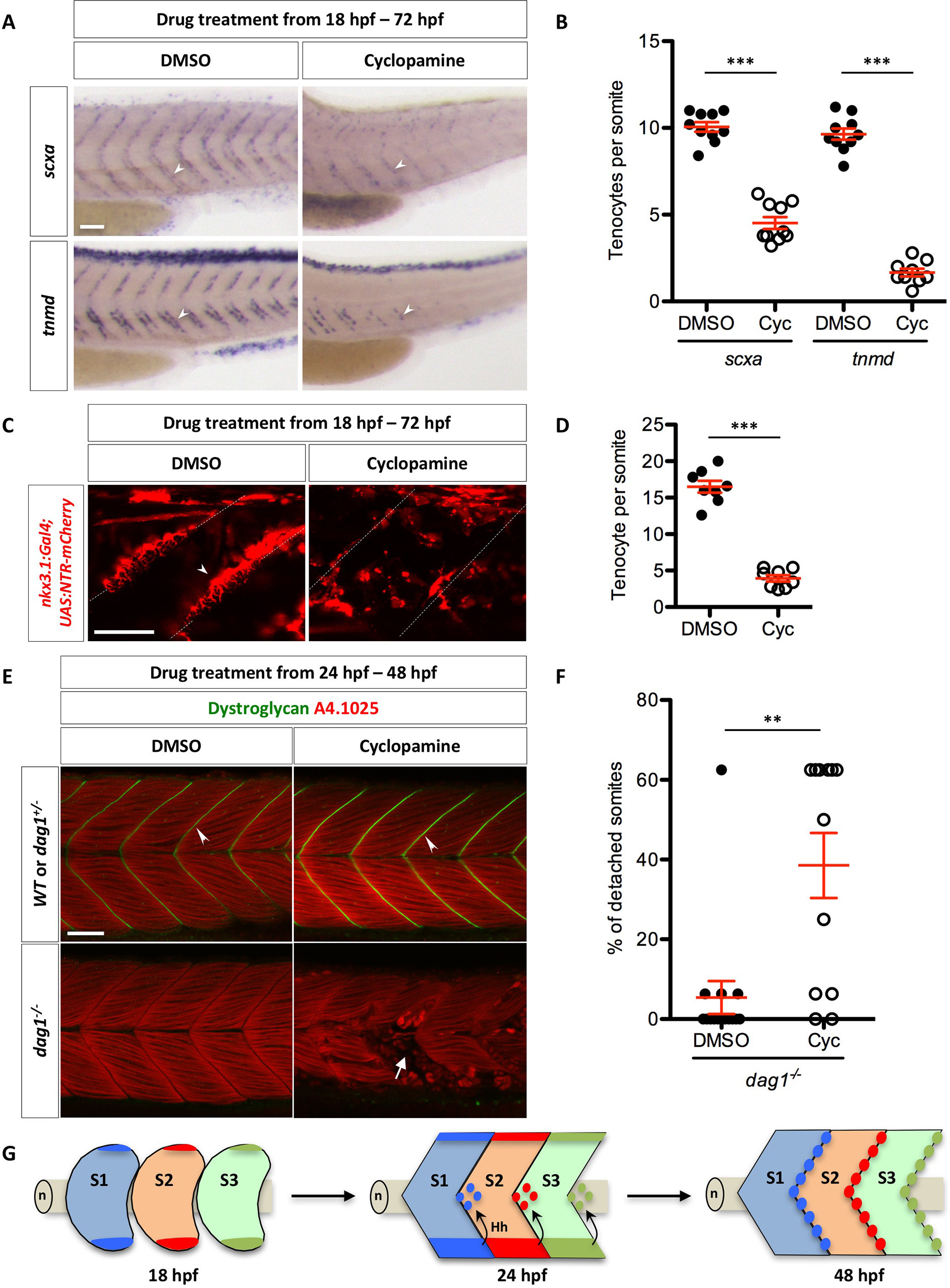
Regulation of tenocytes by Hh signaling.
(A) Wild-type embryos were treated with DMSO or cyclopamine between 18 hpf and 72 hpf, and stained for the expression of scxa and tnmd. Expression of both scxa and tnmd in tenocytes (arrowheads) were reduced in the presence of cyclopamine compared to DMSO controls. n = 60 embryos per condition. (B) Quantification of the drug experiment in (A). The numbers of scxa+ and tnmd+ tenocytes along each MTJ were scored and averaged between somite 18 and somite 23 in DMSO/cyclopamine treated embryos. Each data point represents the average number from an individual embryo. Cyclopamine-treated embryos had significantly reduced numbers of scxa+ and tnmd+ tenocytes. n = 10 (DMSO/scxa, Cyc/scxa and DMSO/tnmd) and 9 (Cyc/tnmd) embryos. (C) nkx3.1NTR-mCherry embryos were treated with DMSO or cyclopamine between 18 hpf and 72 hpf, and imaged to visualize tenocytes. Reduced number of tenocytes (arrowhead) along the MTJ (dotted lines) was observed in cyclopamine-treated embryos. n = 20 embryos. (D) Quantification of the drug experiment in (C). The number of tenocytes was quantified similarly as in (B). Cyclopamine-treated embryos had significantly reduced numbers of nkx3.1NTR-mCherry tenocytes. n = 8 embryos per condition. (E) Embryos from a cross between two heterozygous dystroglycan fish (dag1+/-) were treated with either DMSO or cyclopamine from 24 hpf to 48 hpf, and stained for Dystroglycan (green) and myosin heavy chain (A4.1025, red). Dystroglycan was present along the MTJ (arrowheads) in sibling controls but absent in dag1-/- fish. Most DMSO-treated dag1-/- fish had normal muscle attachments despite the loss of Dystroglycan. By contrast, cyclopamine-treated dag1-/- embryos had extensive muscle detachment as evident by the presence of coiled-up fibers (arrow). (F) Quantification of muscle detachment in drug treated dag1-/- embryos shown in (E). dag1-/- fish treated with cyclopamine showed a significant increase in the percentage of somites with detached muscles. n = 15 (DMSO) and 12 (Cyc) embryos. Scale bars: 50 μm. (B, D, F) All data are plotted with mean ± SEM indicated. Statistics: Mann-Whitney U test. Asterisks representation: p-value < 0.01 (**) and p-value < 0.001 (***). (G) Model of axial tenocyte formation in zebrafish. At 18 hpf, the dorsal and ventral domains of the sclerotome are induced independent of Hh signaling. By 24 hpf, cells begin to migrate from the ventral sclerotome and generate a population of cells surrounding the notochord (n). Cells always exit from the posterior end of the ventral somite, and eventually occupy an area around the notochord about one half-somite more posterior to the somite of origin. The migration of sclerotome derived notochord associated cells is dependent on Hh signaling. Hh signaling is subsequently required to maintain sclerotomal marker expression in these cells. By 48 hpf, the MTJ is populated by tenocytes derived from the sclerotome from the anterior somite. Three somites (S1, S2, and S3) corresponding to somite 17–19 are depicted in the model.