
- Title
-
In Vivo imaging of molecular interactions at damaged sarcolemma
- Authors
- Roostalu, U., and Strähle, U.
- Source
- Full text @ Dev. Cell

Zebrafish Models for Dysferlinopathy PHENOTYPE:
|
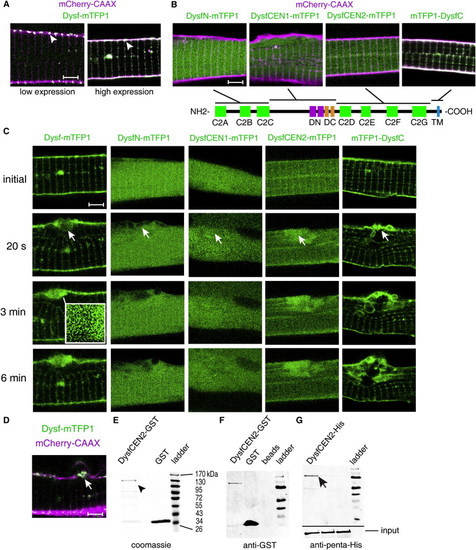
Characterization of Dysf and Its Response to Cell Lesions |
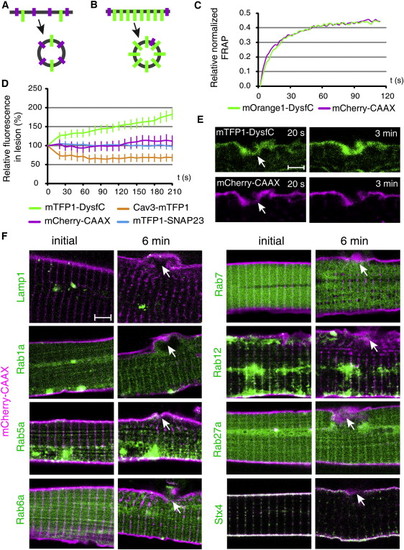
Origin of the Repair Membrane |

Annexin Kinetics in Myofibers |

Distribution of Annexins in the Lesion and the anxa6 Morphant Phenotype PHENOTYPE:
|
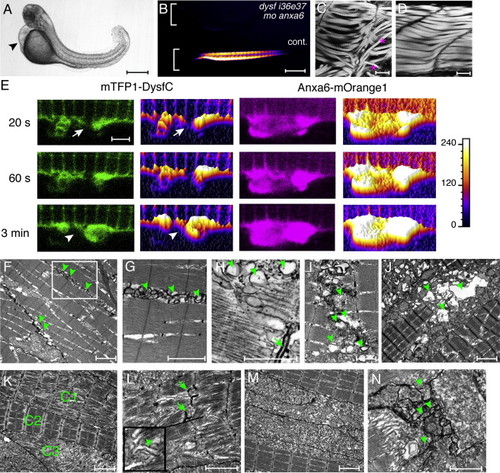
Anxa6/Dysf Double Morphant, Anxa6/Dysf Coexpression, and Ultrastructural Characterization of Muscle Damage PHENOTYPE:
|
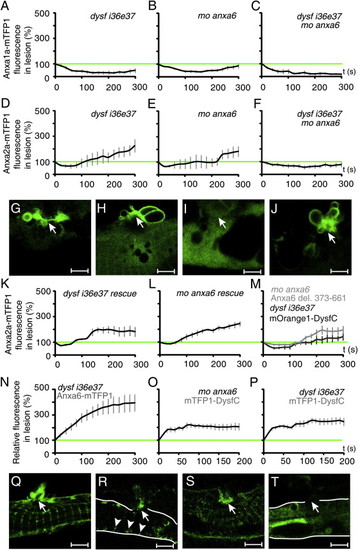
Functional Interactions in Repair Patch Formation |
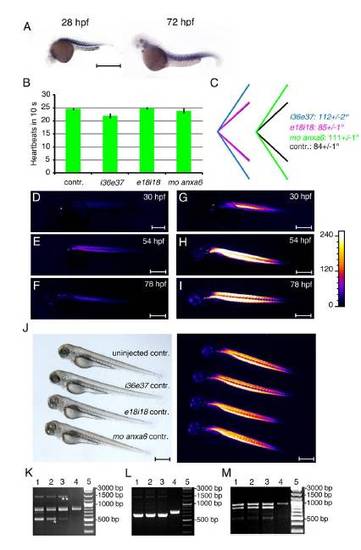
Characterization of dysferlin expression and morpholino-injected embryos. EXPRESSION / LABELING:
PHENOTYPE:
|
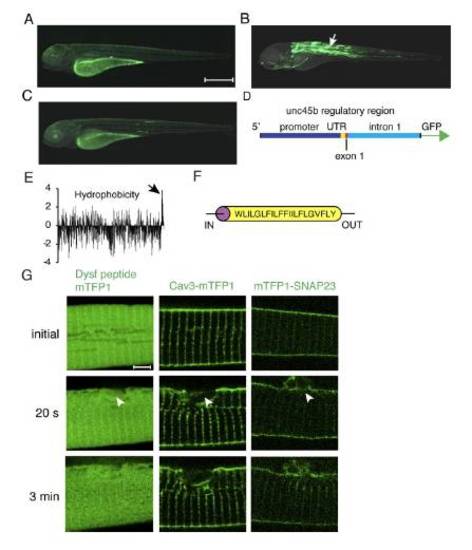
Characterization of the unc45b regulatory region. Dysferlin in silico analysis and membrane protein response to sarcolemmal damage. (A) GFP expression driven by the unc45b promoter from -1799 to -108 relative to the ATG, (B) the unc45b promoter together with the UTR, first exon and the first intron (-1799 to + 1534), (C) the unc45b intron alone upstream of the GFP coding sequence.Schematic illustration of the construct used in (B), ranging from -1799 bp to +1534 bp relative to the unc45b ATG. While the promoter and the intron alone result in weak or no muscle expression (A, C; note the expression in the yolk is unspecific), the full length construct (-1799 to +1534 bp relative to the ATG) of the unc45b gene drives robust expression in the muscle (B, arrow). The first intron of unc45b encompasses a strong transcription enhancer, resulting in specific mosaic expression in skeletal and cardiac muscle cells in transient expression assays. Orientation of embryos: anterior left, dorsal up. (E) In silico analysis of zebrafish dysferlin (Dysf) hydrophobicity (Kyte and Doolittle blot). Positive values on the vertical axis indicate hydrophobic regions. The C-terminus of the protein is towards the right and the arrow points to the transmembrane domain. The majority of Dysf is hydrophilic and the hydrophobic transmembrane domain is followed by a 22 amino acid long hydrophilic stretch at the very C-terminus. (F) Dysf transmembrane domain prediction. (G) No accumulation of Dysf C-terminal 22 amino acid peptide, caveolin 3 (Cav3-mTFP1) or mTFP1-SNAP23 (Soluble NSF Attachment Protein 23) could be observed in ruptured (arrowhead) sarcolemma. Scale bar: A-C, 500 μm; G, 4 μm. |

Damage response of vesicle markers. (A) Lysosome-associated protein transmembrane 4a (Laptm4a) or lysosome-associated membrane protein 2 (Lamp2), fused to mTFP1 in their C-terminus (green) were co-expressed with prenylated mCherry (magenta). Cells were damaged (arrow) with the two photon laser and localization of lysosomal markers was recorded. No large-scale redistribution of lysosomes was evident, although limited amount of mTFP1 could occasionally be observed near the lesion. (B-C) Quantification of lysosome dynamics at the site of lesion. Fluorescence change (%) relative to 20 s time point after injury is depicted. (D) Lysosomes (arrowhead) were observed near the lesion (arrow) only when the rupture occurred close to them. Even then, they did not relocate and fuse to the membrane. (E) Quantification of various endocytosis or exocytosis markers at the site of sarcolemmal lesion. The vertical axis demonstrates fluorescence change at the site of lesion (%) relative to the 20 s time point. Due to the irregular distribution of the vesicles across the cell, the analysis was carried out relative to the 20 s time point. By this time mTFP1-DysfC had already significantly increased at the site of lesion, yet still continued to accumulate also afterwards. Such clear trend could not be observed for any of the studied vesicle markers. Scale bars: A, 4 μm; D, 2 μm |

Expression and subcellular localization of annexins in zebrafish. (A) In situ hybridization of annexins anxa1a, anxa1b, anxa2a, anxa6, anxa11a and anxa11b was carried out at 72 hpf (hours post-fertilization). Lateral view of the trunk region is shown. Expression of anxa1a and anxa2a is detectable in the muscle (mRNA enriched at the myoseptal regions) and in the periderm. Anxa6 is highly expressed in the muscle tissue, whereas by 72 hpf only limited amount of anxa11a could be detected in the muscle. Anxa1b and anxa11b expression are limited to the periderm. (B) Reverse transcriptase-PCR was carried out on adult zebrafish skeletal muscle (lanes 1-7) and dermis (lanes 8-14) samples: anxa1a (lanes 1, 8), anxa1b (lanes 2, 9), anxa2a (lanes 3, 10), anxa2b (lanes 4, 11), anxa6 (lanes 5, 12), anxa11a (lanes 6, 13) and anxa11b (7, 14). DNA ladder: 15. Anxa1a, anxa2a and anxa6 are abundant in adult zebrafish muscle tissue. (C) In addition to cytoplasmic localization, strong nuclear signal (asterisk) is visible for Anxa1a and Anxa11a, when fused with mTFP1 (green). Weaker nuclear accumulation is evident for Anxa6-mTFP1 and no nuclear signal could be detected for Anxa2a-mTFP1. mCherry-CAAX marks the cell membrane in magenta. (D) Translocation of Anxa6-mTFP1 to the cell membrane in paraformaldehyde fixed animals. (E) Annexins tagged with mTFP1 (green) accumulate in the sarcolemmal lesion after injury (arrow). Larger cellular wounds eventually lead to the binding of annexins to the membrane distant from the site of injury (arrowhead). (F) Small sarcolemmal wounds lead to temporary accumulation of Anxa1a at the lesion. (G) Large wounds result in persistent Anxa1a localization at the plasma membrane. (H) The relative accumulation kinetics of the studied annexins does not depend on lesion size. Anxa6-mOrange1 (purple) accumulates faster in the large lesion than Anxa2a-mTFP1 (green). Similar results were obtained for small lesions (compare to Figure 4A). (I) Accumulation of annexins does not depend on the expression level. Two cells with 30% difference (largest difference in the study) of Anxa1a-mTFP1 expression were injured. In none of them did the accumulation take place at 60 s after rupture. Scale bars: A, 185 μm; C-F, H-I, 4 μm; G, 65 μm. EXPRESSION / LABELING:
|

Annexin damage response. (A-H) Anxa2a-mTFP1 at the sarcolemmal rupture (arrow) at 3 min time point after damage. Anxa2a-mTFP1 response to sarcolemmal lesion was unperturbed in dysf control morpholino-injected embryos (A, D) and in anxa6 control morpholino-injected embryos (B, E). (F) Dysf-mOrange1 rescues Anxa2a-mTFP1 repair patch morphology in dysf i36e37 morphants. (G) Anxa6-mOrange1 rescues Anxa2a-mTFP1 repair patch morphology in anxa6 morphants. (C, H) Unaltered Anxa2a-mTFP1 accumulation and patch formation in dysf e18i18 morphants. Scale bar: A-D, F-G, K-O, 4 μm; E, 65 μm. |
Reprinted from Developmental Cell, 22(3), Roostalu, U., and Strähle, U., In Vivo imaging of molecular interactions at damaged sarcolemma, 515-529, Copyright (2012) with permission from Elsevier. Full text @ Dev. Cell