
- Title
-
The development of lateral line placodes: taking a broader view
- Authors
- Piotrowski, T., Baker, C.V.
- Source
- Full text @ Dev. Biol.
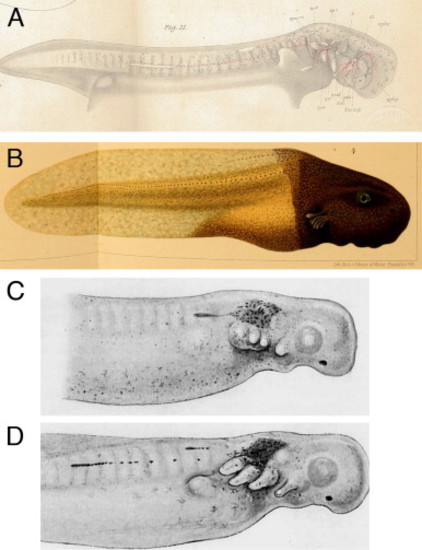
In the majority of non-cartilaginous vertebrates, the posterior lateral line placodes migrate and deposit lateral line sense organs. (A) The posterior lateral line placode often splits into a dorsal and a ventral component, as seen in the mudpuppy Necturus (an aquatic salamander) (reproduced from Platt, 1896). (B) Fusion of the trunk of a lightly pigmented frog embryo (Rana palustris) with the head of a darkly pigmented embryo (Rana sylvatica) demonstrated that pigmented lateral line placodes from the head migrate onto the trunk where they deposit sense organs ( Harrison, 1904). (C,D) Stone (1933) transplanted pieces of Nile blue-stained donor ectoderm containing the posterior lateral line placode onto an unstained spotted salamander (Ambystoma maculatum; Ambystoma punctatum) embryo. (C) 2 days post-transplantation the stained club-shaped placode had started to migrate. (D) By 3 days post-transplantation, the placodes had migrated further and deposited sense organs |
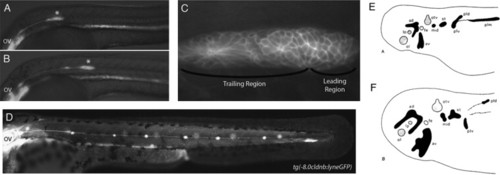
The posterior lateral line placode migrates; other lateral line placodes elongate to form sensory ridges. (A) At 24 hpf the posterior lateral line primordium (asterisk) in a zebrafish embryo has fully migrated onto the somites from its origin near the otic vesicle (OV). (B) The primordium has recently deposited the first proneuromast. (C) Higher magnification of the primordium in (A). Rosettes begin to form a short distance away from the leading region within the migrating primordium. (D) The post-embryonic lateral line is composed of 4–6 proneuromasts in a row along the horizontal myoseptum. Panels A–D are reproduced from Aman and Piotrowski (2011) with permission from Cell Adhesion and Migration. (E,F) Camera lucida drawings of flat mounts of the head ectoderm of axolotl embryos at (E) stage 25 and (F) stage 37, showing lateral line placodes in black and other placodes in gray. Panels E and F reproduced from Northcutt et al. (1994) with permission from Wiley and Sons. Abbreviations: ad, anterodorsal lateral line placode; av, anterodorsal lateral line placode; lp, lens placode; mid, middle lateral line placode; ol, olfactory placode; otv, otic vesicle; pld, dorsal subdivision of posterior lateral line placode; plm, main subdivision of posterior lateral line placode; plv, ventral subdivision of subdivision of posterior lateral line placode; st, supratemporal lateral line placode. |
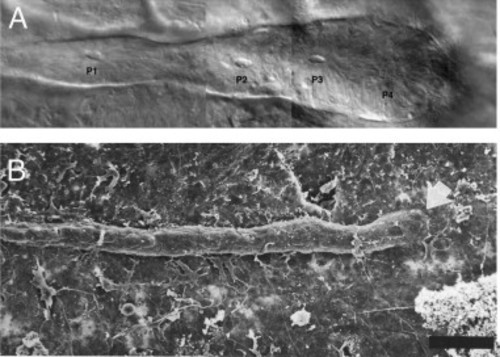
Posterior line placodes vary in shape. (A) Nomarski brightfield image of a posterior lateral line primordium in a blind Mexican tetra (blind cavefish, Astyanax fasciatus). The primordium is club-shaped and sensory organs begin to form within the migrating primordium (P1–P3) (reproduced from Sapède et al., 2002 with permission from Development). (B) Scanning electron micrograph of the posterior lateral line placode in an axolotl larva (Ambystoma mexicanum). The placode is much thinner than in teleosts. Image reproduced with permission from the International Journal of Developmental Biology ( Smith, 1996). Scale bar 100 µm. |
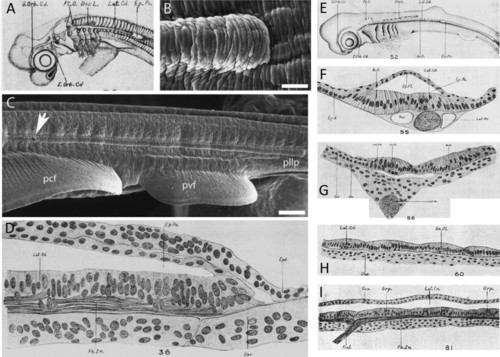
In cartilaginous fishes, an epidermal pocket or tunnel forms above the posterior lateral line primordium as it elongates. (A) Lateral surface view of a 20 mm spiny dogfish (Squalus acanthias) specimen. The elongating posterior lateral line primordium (lateral sensory cord/ridge, Lat.Cd.) and epidermal pocket or tunnel (Ep.Po.) are clearly visible (sketch reproduced from Johnson, 1917). (B) Scanning electron micrograph of the advancing tip of the posterior lateral line primordium within the epidermal pocket/tunnel in the little skate Leucoraja erinacea (reproduced from Gillis et al., 2012). (C) The sensory ridge extends caudally along the entire length of the trunk. The opening to the epidermal tunnel is indicated by a white arrow (reproduced from Gillis et al., 2012). (D) Horizontal section through the growing end of the posterior lateral line primordium in a 15 mm S. acanthias specimen, showing the double-layered epidermal pocket/tunnel (Ep.Po) forming dorsal to it (modified from Johnson, 1917). (E) Lateral surface view of a 27 mm S. acanthias specimen (reproduced from Johnson, 1917). (F) Transverse section through a 27 mm S. acanthias specimen at a level caudal to the first dorsal fin, where the lateral sensory ridge is still covered by the superficial epithelial pocket (reproduced from Johnson, 1917). (G) Transverse section through a 27 mm S. acanthias specimen at a level rostral to the epidermal pocket/tunnel, where the lateral sensory ridge is exposed to the surface (reproduced from Johnson, 1917). (H) Horizontal section through a 29 mm S. acanthias specimen. The hair cells are arranged into small groups (reproduced from Johnson, 1917). (I) Longitudinal section of the lateral canal in a S. acanthias pup. The sketch shows the general histological morphology of the sensory epithelium, which is now enclosed within an epithelial canal (Lat.Cn.), and its innervation (reproduced from Johnson, 1917). Abbreviations: Ac.O., accessory organ; Can., canal wall; Der, dermis; Dor.L., anlage of dorsal line of pit organs; Ep.Fl., epidermal fold; Ep.Po., epidermal pocket or tunnel; Epd., epidermis; Fb.Zn., longitudinal nerve fiber zone; Grp., group of hair cells; I.Orb.Cd., supraorbital sensory cord; Lat.Cd., lateral sensory cord; Lat.Cn., lateral canal; Lat.Nv., lateral line nerve; pcf, pectoral fin; pllp, posterior lateral line primordium; Pt.O., anlage of isolated pit organs; pvf, pelvic fin; Rml., ramulus lateralis (little branch of lateral line nerve); S.Orb.Cd., supraorbital sensory cord; Sn.Cl., secondary sense cell; Vas., vascular space. |
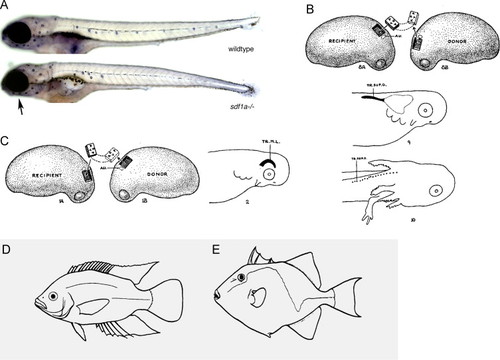
(A) Alkaline phosphatase staining reveals that in sdf1a mutant zebrafish, the posterior lateral line primordium has not migrated all the way to the tail tip. However, the anterior lateral line neuromasts have formed normally around the eye and on the lower jaw (black arrow). Images from Mark Lush and T.P. (B) Stone (1928b) transplanted pre-otic ectoderm from a donor spotted salamander (Ambystoma maculatum; A. punctatum) embryo (on the right of the diagram) into the position of the posterior lateral line placode of the host embryo (on the left of the diagram). The placode migrated or extended and gave rise to a defined number of neuromasts (bottom image, embryo with gills and forelimb) (reproduced from Stone, 1928b). (C) Reciprocally, Stone (1928b) transplanted post-otic ectoderm into the position of the pre-otic supraorbital lateral line primordium and reversed its orientation by 180 degrees. The transplanted placode migrated or extended around the eye and differentiated into neuromasts (reproduced from Stone, 1928b). (D,E) The morphology of the lateral line is diverse between different species of teleosts (reproduced from Coombs et al., 1988 with permission from Springer Verlag). (D) Cichlid with an interrupted trunk line. (E) Triggerfish with a zig-zag shaped lateral line. |
Reprinted from Developmental Biology, 389, Piotrowski, T., Baker, C.V., The development of lateral line placodes: taking a broader view, 68-81, Copyright (2014) with permission from Elsevier. Full text @ Dev. Biol.