
- Title
-
Analysis of a zebrafish dync1h1 mutant reveals multiple functions for cytoplasmic dynein 1 during retinal photoreceptor development
- Authors
- Insinna, C., Baye, L.M., Amsterdam, A., Besharse, J.C., and Link, B.A.
- Source
- Full text @ Neural Dev.
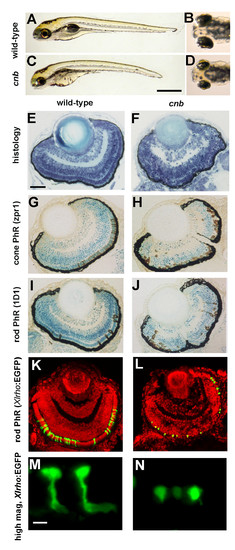
The cnb mutation disrupts photoreceptor development. (A-D) Side (A, C) and dorsal (B, D) views of 5-dpf wild-type (A, B) and cnb mutant (C, D) embryos. (E-F) Retinal histology of wild-type (E) and cnb (F) retina at 4 dpf. (G, H) Zpr-1 immunoreactivity (brown) to label double cone photoreceptors (PhR) in 4-dpf wild-type (G) and cnb embryos (H). (I, J) ID1 immunoreactivity (brown) to label rod photoreceptors in 4-dpf wild-type (G) and cnb embryos (H). Nuclei in (G, H) are counterstained with methylene blue. (K-N) Confocal microscopy of wild-type (K, M) and cnb (L, N) whole retina (K, L) and individual rod photoreceptor cells (M, N) from Tg(Xlrho:EGFP)fl1 fish. Green, Xlrho:EGFP-positive cells; red, propidium iodide positive nuclei. Scale bars: 1 mm in (A, C); 40 μm in (E-L); 5 μm in (M, N). EGFP, enhanced green fluorescent protein. |
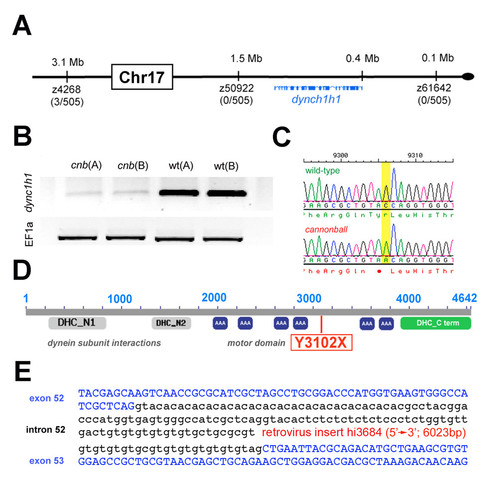
The cnb locus encodes dync1h1. (A) Map of the cnb critical genomic region and exon/intron representation of dync1h1. Linked microsatellite markers and recombinants are placed onto the ZV7 zebrafish genomic map. (B) RT-PCR results from two independent cDNA samples each of cnb or wild-type embryos. Top image of bands from reactions with dync1h1 primers and bottom image of bands from reactions with EF1alpha primers. (C) Chromatograms showing wild-type (top) and cnb sequence flanking the C-to-A mutation. (D) Schematic of the Dync1h1 protein and location of the nonsense mutation. (E) Location of retrovirus insertion for dync1h1 mutant allele hi3684. |
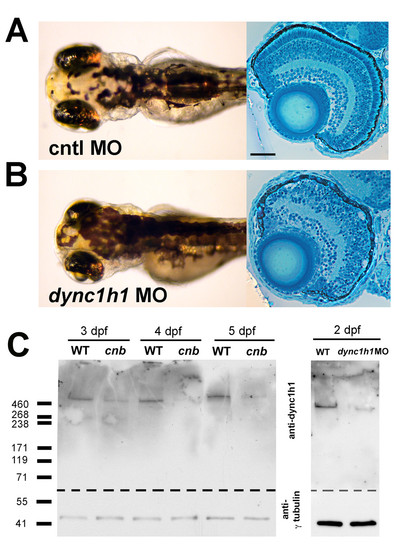
Morpholino knock-down of dync1h1 phenocopies cnb mutants. (A, B) Dorsal views and retinal histology of control (cntl) morpholino (MO) injected (A) and dync1h1 ATG MO injected (B) embryos shown at 3.5 dpf. Note the smaller eyes, expanded melanophores, and photoreceptor dysmorphogenesis. (C) Western blot of wild-type (WT) versus cnb embryos at 3, 4, and 5 dpf (left blot) and wild-type and dync1h1 morphants at 2 dpf (right blot). Each blot was cut and probed with either antisera against human DYNC1H1 (amino-terminal 321 amino acids; top) or gammaTubulin (bottom). Scale bar: 40 μm for retinal histology. EXPRESSION / LABELING:
PHENOTYPE:
|
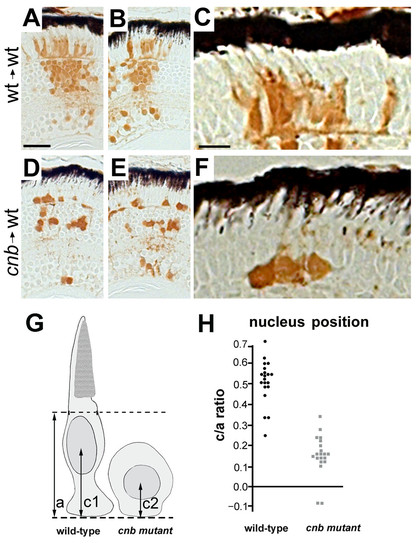
Photoreceptors from cnb/dync1h1 mutants show cell-autonomous defects in morphogenes. (A-F) Images of histology sections from genetically mosaic retinas of wild-type (wt) donor, wild-type host (A-C) or cnb donor, wild-type host (D-F). Two examples are shown at low magnification to indicate overall clone size and one example at higher magnification to show photoreceptor morphology. Donor cells are label with the brown lineage tracer. Black retinal pigment epithelium cells provide reference for orientation. Note the lack of elongated morphology in cnb photoreceptor cells. Scale bars: 40 μm in (A, B, D, E); 15 μm in (C, F). (G) Schematic showing how nuclear position was evaluated. Dashed lines represent the external limiting membrane (upper) and apical side of the outer plexiform layer (bottom) for the host retina. (H) Scatter plot of nuclear position (c/a ratio) for donor cells of wild-type (black, left side) or cnb (grey, right side) genotypes. |
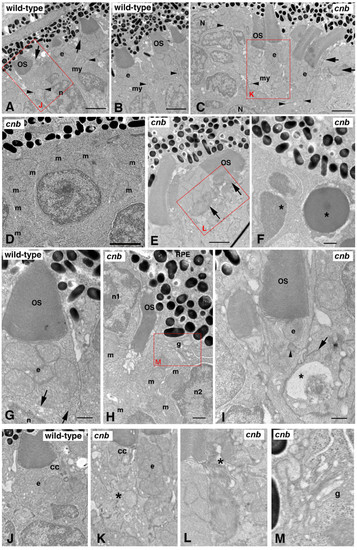
The cnb mutation disrupts photoreceptor polarity and outer segment formation. (A, B) TEM of a wild-type central retina at 3 dpf showing normal structure of outer segments (OS) with aligned discs adjacent to the ellipsoid (e) region of the inner segments with bundles of tightly compacted mitochondria. The myoid (my) of the inner segment is the region between the mitochondria-rich ellipsoid and the elongated nuclei (n). Dense adherens junctions (arrowheads) can be seen at the external limiting membrane. Arrows in (A) point to connecting cilia. See also (J) for high magnification. (C-F, H, I) TEM of cnb mutant retina. (C) TEM of a cnb central retina at 3 dpf with both normal photoreceptor outer segments (left) and cells with disorganized outer segments (right, arrows). Adherens junctions are indicated by arrowheads. Overall, photoreceptors in the peripheral retina showed more severe defects, including cells that failed to form a polarized inner segment and cells with pyknotic nuclei. (D) Higher magnification of peripheral cnb photoreceptors that lack polarity and display dispersed mitochondria (m). (E) Normal cnb photoreceptors (left) next to an abnormal cell with a disrupted outer segment (arrows). See (L) for higher magnification. (F) Photoreceptors at the periphery with pyknotic nuclei (asterisks) and dense granular cytoplasm. (G) High magnification image of polarized wild-type cone photoreceptor at 3 dpf showing normal outer segment, typical ellipsoid with clustered mitochondria, and a narrow myoid with membrane vesicles and cisternae (arrows). (H, I) Central cone photoreceptors of cnb mutant eyes showing abnormal polarity. In (H), the cell on the left labeled n1 has its ellipsoidal mitochondria (m) on the opposite side of the nucleus from the retinal pigment epithelium (RPE) with an adjacent miss-aligned outer segment. The cell on the right (n2) has its Golgi complex (g) adjacent to RPE and the mitochondria (m) is scattered throughout the cytoplasm. A higher magnification of the Golgi is shown in (L). In (I), a cnb cone photoreceptor with a normal OS has a single mitochondrion at the position of the ellipsoid (e) and a displaced centriole (arrowhead). The adjacent cell on the right displays an abnormally positioned synaptic region with a large vacuole (asterisk) and a synaptic ribbon (arrow) adjacent to the inner segment of the cone cell in the middle. (J-M) Higher magnification insets of the red boxed regions highlighting the connecting cilium (cc) and ellipsoid (e) regions. Asterisk in (K) indicates a cell with a disorganized inner segment adjacent to a cell with a more normal ellipsoid. Asterisk in (L) indicates disorganized outer segment. Scale bars: 2 μm in (A-C); 0.5 μm in (F-I). |
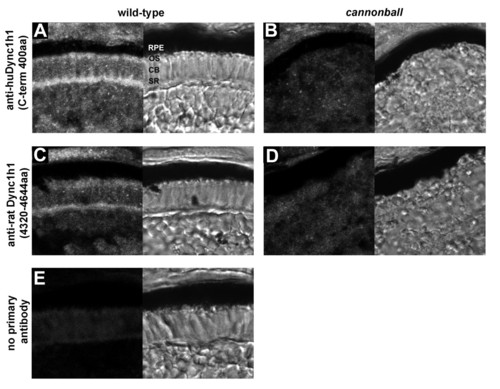
Immunolocalization of Dync1h1 in developing photoreceptors. (A-E) Cryosections of the photoreceptor layer from wild-type retina (A, C, E) or cnb mutant retina (B, D) each at 4 dpf. Antisera directed against two different regions of Dync1h1 was used: (A, B) anti-human DYNC1H1 (huDYNC1H1; carboxy-terminal 400 amino acids) and (C, D) anti-rat Dync1h1 (amino acids 4,320 to 4,644). Primary antibodies were omitted in (E). For each panel, the fluorescent image is shown on the left and the transmitted light image on the right. CB, cell body; OS, outer segment; RPE, retina pigment epithelium; SR, synaptic region. |
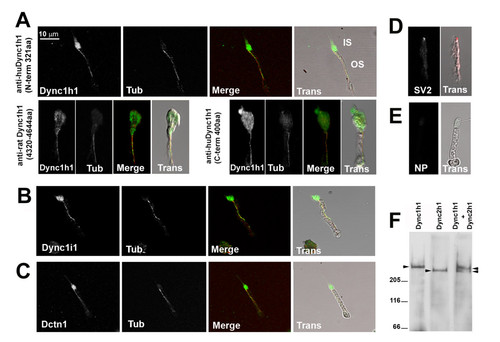
Dync1h1, Dync1i1, and Dnct1 immunoreactivity is associated with photoreceptor axonemes. Immunocytochemical localization of proteins in isolated adult zebrafish photoreceptor cells. (A) Immunocytochemical localization of Dynein1 heavy chain (Dync1h1) using antisera directed against three different epitopes. (B) Immunocytochemical localization of Dynein1 intermediate chain (Dync1i1), (C) Dynactin1/p150 (Dctn1), and (D) Synaptic vesicle protein 2 (SV2). (E) Immunostaining in which the primary antibody was omitted. In (A-C), cells were co-stained with acetylated α-tubulin (Tub) to highlight the axoneme. Color merged (Merge) and transmitted light images overlaid with Merge images (Trans) are shown for each panel. IS, inner segment; OS, outer segment. NP, no primary antibody. (F) Western blot analysis on detergent-extracted photoreceptor cytoskeleton (DEPC) fraction shows the specificity of the Dync1h1 antibody, as the Dync2 h2 antibody recognizes a discrete and faster migrating band. EXPRESSION / LABELING:
|
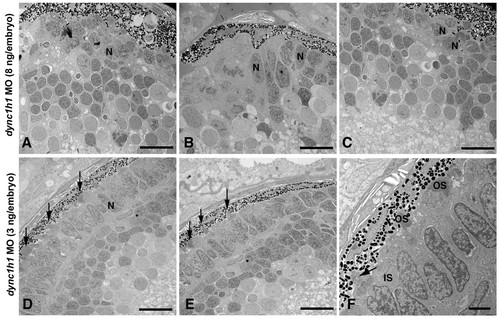
Knock-down of Dync1h1 phenocopies cnb mutants. (A-C) TEM of retinas from 3-dpf embryos injected with a high dose (8 ng/embryo) of dync1h1 morpholino (MO). Photoreceptors across the entire retina failed to form outer segments and had rounded nuclei (N). (D-L) TEM views of embryos injected with a lower amount (3 ng/embryo) of dyn1 hc1 morpholino. Outer segments formed in some photoreceptors at the central (D, E) and peripheral retina (F) (arrows). Most lower dose morphant cells still failed to elongate outer segments. Scale bars: 10 μm in (A-E); 2 μm in (F). PHENOTYPE:
|
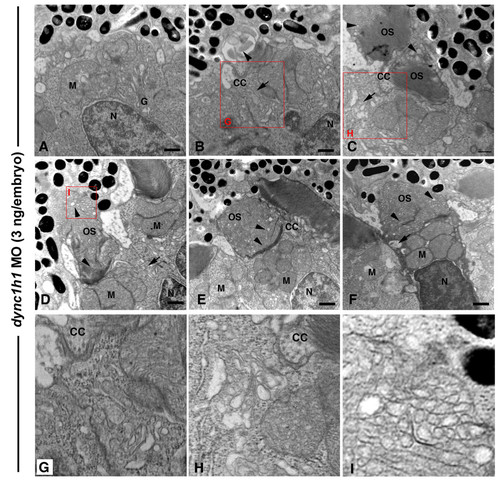
Dync1h1 knock-down causes dose-sensitive outer segment defects. TEM of retinas from 3-dpf embryos injected with a high dose (8 ng/embryo) of dync1h1 morpholino (MO). (A) A cone cell with Golgi apparatus (G) localized in the central ellipsoid region and mitochondria (M) located near the nucleus (N). (B-I) Cone cells with accumulation of vesicles (arrows) at the base of the connecting cilium (CC) and presence of disorganized disc structures within the outer segment (OS; arrowheads). (D-F) Severely disrupted outer segments showing vesiculation and tubulation of discs structures distally (arrowheads), yet normal discs at the base. Large vacuoles and vesicle accumulations were found at the base of the connecting cilium (arrows). The condensed cell in (F) is consistent with the morphology of early stage apoptosis. (G-I) Higher magnification insets of the red boxed regions. Scale bars: 0.5 μm in (A-F). PHENOTYPE:
|
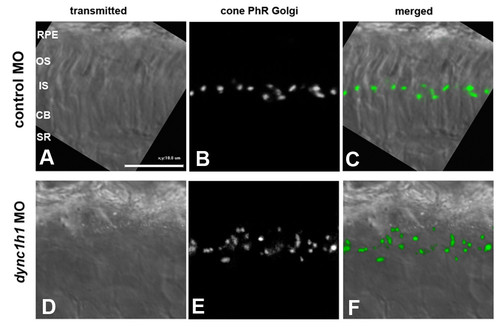
Localization of Golgi apparatus in dync1h1 morphants. (A-F) Expression of a tol2-based gnat2:Man2a(1-100aa)-GFP construct in either control morphants (3 ng/embryo) (A-C) or dync1h1 morphants (3 ng/embryo) (D-F). Cryosections of the photoreceptor (PhR) layer at 4 dpf is shown. Embryos were treated with phenylthiourea to block pigment synthesis. Note that disorganization and mis-positioning of the Golgi apparatus in dync1h1 morphants. CB, cell body; IS, inner segment; MO, morpholino; OS, outer segment; RPE, retina pigment epithelium; SR, synaptic region. PHENOTYPE:
|
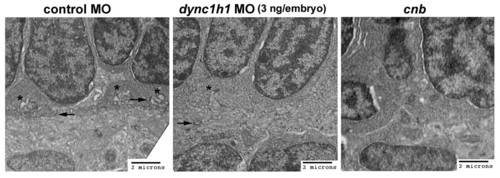
Ultrastructure of synaptic region in cnb mutants and dync1h1 morphants. TEMs of the photoreceptor synaptic region and outer plexiform layer of 3-dpf embryos. (A) Control morphants (3 ng/embryo) showed well developed synaptic input invaginations (asterisks) and numerous floating ribbons (arrows). (B) dync1h1 morphants (3 ng/embryo) had less defined synaptic input invaginations (asterisk) and significantly reduced number of floating ribbons (arrow). (C) cnb mutants did not show synaptic input invaginations and typically lacked ribbons. MO, morpholino. PHENOTYPE:
|
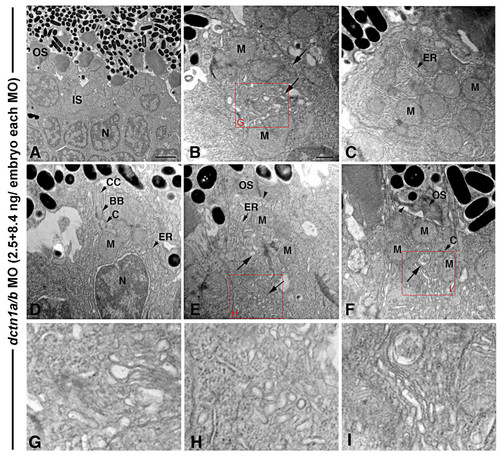
Double dnct1a/b knock-down primarily affects the inner segment. (A) TEM view of the central retina of a dctn1a/b morphant at 3 days showing normal, albeit short, outer segment structures. (B-F) TEM views of individual photoreceptors at the peripheral dctn1a/b morphant retina with inner segment polarization defects, including a dispersion of mitochondria (M) within that region (B, C, F), lack of a distinct myoid region (D) and positioning of the ER/Golgi complex among dispersed mitochondria (arrows in (B, E, F)). In addition, accumulation of Golgi associated vesicles in the inner segment (arrow in (B, E, F)) and shortened outer segment structures (arrowheads) were also seen (E, F). (G-I) Higher magnification insets of the red boxed regions. For morpholino (MO) injections, dnct1a splice MO was used at 2.5 ng/embryo, while the dnc1b splice MO was used at 8.4 ng/embryo (for a total MO concentration of 10.9 ng/embryo). BB, basal body; C, centriole; CC, connecting cilium; ER; endoplasmic reticulum; IS, inner segment; M, mitochondria; OS, outer segment. Scale bar: 2 μm in (A); 0.5 μm in (B-F). PHENOTYPE:
|
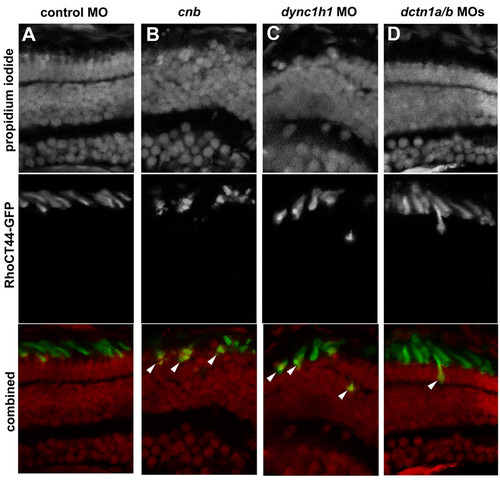
Efficient transport of Rhodopsin(CT44)-GFP to the outer segment is disrupted with loss of Dync1h1 or Dctn1. Cryosections of 3.5-dpf retina from transgenic Tg(1.3xops:xRhoCT44-GFP) embryos in which the carboxy-terminal 44 amino acids of Rhodopsin are fused with GFP. Localization of GFP was evaluated in photoreceptors from (A) control morphants, (B) cnb mutants, (C) dync1h1 morphants or (D) dctn1a/b morphants. Upper row shows propidium iodide staining to highlight nuclei. Middle row shows RhoCT44-GFP. Bottom row shows color combined images. Arrowheads indicate cells with mislocalized GFP to the cell body region. EXPRESSION / LABELING:
PHENOTYPE:
|
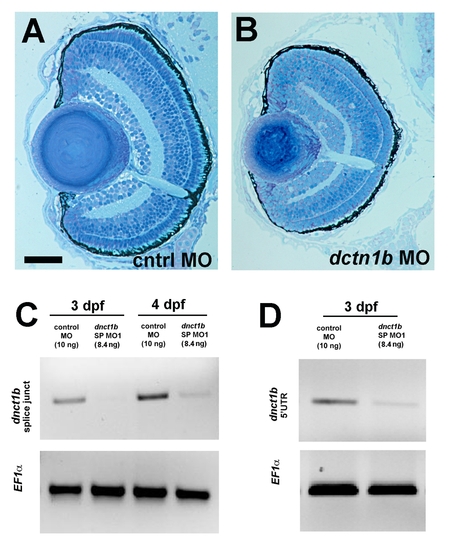
Morpholino knock-down of dnct1b. (A, B) Ocular histology of control (A) and dnct1b (B) morphants at 3 dpf. (C) RT-PCR analysis of dnct1b morpholino-injected and control morpholino-injected embryos indicating efficient targeting of the transcript. Primers in the top image bind to regions flanking the morpholino-targeted splice junction. (D) RT-PCR analysis of dnct1b and control morphants. Primers in the top image bind to the 5′ untranslated region of dnct1b. Bottom images in (C, D) show results using control EF1alpha primers that indicate the dnct1b morpholino destabilizes the targeted mRNA. The age of embryos used to make cDNA is shown at the top of (C, D). Scale bar: 40 μm in (A, B). |