
- Title
-
Modulation of BMP activity in dorsal-ventral pattern formation by the Chordin and Ogon antagonists
- Authors
- Wagner, D.S., and Mullins, M.C.
- Source
- Full text @ Dev. Biol.
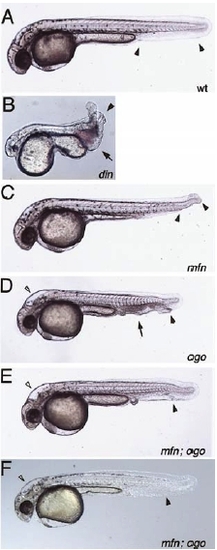
mini fin (mfn) double mutant analysis. Phenotypes observed in embryos from crosses between mfn;din double heterozygous fish. (A) In wild type embryos a single ventral tail fin extends the full length of the tail (arrowheads). (B) The din mutant displays multiple ventral tail fins (arrowhead), an edema posterior to the yolk extension at the site of blood formation (arrow) and a variably reduced head and eyes. (C) mfn mutant embryos display a gap in the ventral tail fin (arrowheads). Additional phenotypes to (A and C) observed in embryos from crosses of mfn;ogon double heterozygous fish (D and E). (D) The ogon mutant is characterized by multiple ventral tail fins (black arrowhead), an edema posterior to the yolk extension (arrow) and an altered head morphology (white arrowhead). (E) Embryos exhibiting the ogon head phenotype (white arrowhead) and a much less severe or wild type tail phenotype (black arrow head). This embryo had only a small edema posterior to the yolk extension without the pooling of blood or the multiplicated ventral tail fins seen in D. |

Loss of chordino has no apparent effect on gene expression in swirl (swr) and snh embryos. msxB expression at the 5-somite stage (A–D). (A) Wild type. (B) din embryos exhibit a variable reduction of anterior msxB staining in the presumptive neural crest (nc). (C and D) A similar small range of msxB expression in an anterior ventral position (arrowhead) is observed in embryos from both swirl single heterozygous and swirl;din double heterozygous crosses: shown here are weak (C) and strong (D) phenotypes. Expression of foxd3 (formerly fkd6) in the neural crest progenitors in 5-somite stage snh mutants is not affected by loss of din (E–H, lateral views, dorsal to the right; insets are dorsal-anterior views with anterior to the left). (E) Expression of foxd3 in the prospective cranial neural crest (bracket) and pax2.1 in the mid/hindbrain boundary (white box in inset) of a wild type embryo. (F) din mutant embryos exhibit a variable decrease in the width of anterior neural tissue as seen by decreased spacing between the domains of foxd3 expression and the width of the mid/hindbrain boundary. (G and H) The range of expanded foxd3 expressing neural crest phenotypes observed in snhty68a embryos. The degree of expansion was unaffected by din genotype; both embryos shown are homozygous mutant for din as well as snhty68a. Classification of the embryos as either strong, moderate or weak snh mutant phenotypes (G is weak, H is strong), followed by examination of the din genotype revealed the following: strong, 4 din/din, 4 din/+; moderate, 3 din/din, 3 din/+, 4 +/+; weak 2 din/din, 3 +/+. In crosses with ogon, 1 of 5 embryos with a strong snh phenotype was homozygous mutant for ogon. Of 4 embryos with a weak snh phenotype, none was homozygous mutant for ogon (data not shown). EXPRESSION / LABELING:
|

swirl and snailhouse double mutant analysis. Phenotypes observed in embryos from crosses between swirl; din double heterozygous fish (A–F). (A) Wild type embryo at the 5-somite stage (dorsal to the right, anterior up). (B) A swirl mutant embryo at the 5-somite stage. Morphogenetic movements have driven the somitic tissue (arrowhead) and the tail bud off the yolk. These mutants die by the 14-somite stage (~ 16 hpf). (C) Wild type embryo at 1 dpf displaying a single ventral tail fin (arrowhead). (D) The weakly dorsalized, dominant swirl phenotype is characterized by a partial loss of the ventral tail fin (absent between the arrowheads). (E) din mutant embryos have multiple ventral tail fins (arrowhead) and a large edema posterior to the yolk extension (arrow). (F) The dominant dorsalized swirl mutant phenotype is epistatic to din, as evident by the absence of the ventral tail fin (arrowheads) in this din-/- ;swr+/- embryo. However some aspects of ventralization are still present, such as a minor edema (arrow) and a multiplicated ventral tail fin tip (arrowhead in the inset, which is a slightly different view). For snh double mutant analysis, 3 classes of 1 dpf phenotypes were observed in embryos from crosses between snh; din double heterozygous fish; wild type embryos as in A, din mutant embryos as in E and snh mutant embryos (G). snh embryos are truncated posteriorly and the trunk tissue is twisted above the yolk (arrowhead). Phenotypes observed in embryos from crosses of swirl;ogon double heterozygous fish (H and I). Wild type and swirl mutant embryos were observed identical to those in A, B, and C. (H) ogon mutants at 1 dpf exhibit multiple ventral tail fins (arrowhead) and an edema posterior to the yolk extension (arrow) as well as a swelling of the hindbrain ventricle (white arrowhead). (I) The dominant dorsalized swirl phenotype is epistatic to ogon as evidenced by embryos with a gap in the ventral tail fin (arrow), which also display an ogon head phenotype (white arrowhead) and multiplicated ventral tail fin tip (black arrowhead). |

Loss of ventral tail fin in din mutants. (A) Wild type embryos at 1dpf have a single ventral tail fin (arrows in ventral view inset). (B) A strong dintt250 mutant embryo at 1 dpf displays multiple ventral tail fins (white arrowheads). (C) Occasional dintt250 embryos display gaps in the multiplicated ventral fin (gap, black arrowheads; ventral fins, white arrowheads). dinm52 embryos also display alterations in the fin structure. (D) Most dinm52 embryos display a full ventral tail fin with minor duplications (white arrowheads) (E) Some embryos display reduced fins (black arrowhead) or gaps, in addition to duplications (white arrowheads). All panels lateral views, ventral view in inset. |

lost-a-fin double mutant analysis. Embryonic phenotypes observed from crosses between laf; din double heterozygous fish (A–C). (A) A wild type embryo at 1 dpf. (B) A typical laf mutant embryo, which lacks the ventral tail fin (arrowheads). din mutant embryos were similar to the embryo in Fig. 3E. (C) A double mutant embryo displaying a sharp dorsal flexure of the tail, no ventral tail fin (arrowheads) and a mass at the posterior end of the yolk extension (arrow). (D) A laf;ogon double mutant exhibiting a sharp dorsal bend in the tail, no ventral tail fin (arrowheads) and a cell mass posterior to the yolk extension (arrowhead). The degree of dorsal flexure and the presence of the cell mass are variable compared to the laf; din embryos. Expression of myoD at the 20-somite stage in embryos from laf;din (E–H) and laf;ogon (I–L) intercrosses. Transverse sections of similar embryos approximately at the level of somite 16 (arrows) are shown as insets (E–H). (E and I) myoD expression in the somitic mesoderm of a wild type embryo. The somite staining does not extend toward the ventral side of the embryo. In contrast to the wild type (E and I), in laf mutant embryos (F) myoD expression in the tail somites extends ventrally beginning at approximately somite 17 (arrowhead). (G) din mutants also exhibited a ventral expansion of myoD expression, but in more posterior somites (arrowhead). (H) The double mutant embryos displayed a characteristic dorsal bend in the tail and ventral expansion of myoD expression at a more anterior somite than in either single mutant (somite 12, arrowhead). (J) laf embryos from laf; ogon crosses displayed a ventral expansion of myoD expression beginning at about somite 19. (K) ogon mutant embryos exhibit a less dramatic expansion of myoD expression in posterior somites at this stage. (L) As with din crosses the laf;ogon double mutants embryos exhibited a ventral expansion of somitic mesoderm at a more anterior position, beginning at about somite 15 (arrowhead). |
Reprinted from Developmental Biology, 245(1), Wagner, D.S., and Mullins, M.C., Modulation of BMP activity in dorsal-ventral pattern formation by the Chordin and Ogon antagonists, 109-123, Copyright (2002) with permission from Elsevier. Full text @ Dev. Biol.