
- Title
-
The zebrafish trilobite gene is essential for tangential migration of branchiomotor neurons
- Authors
- Bingham, S., Higashijima, S.-I., Okamoto, H., and Chandrasekhar, A.
- Source
- Full text @ Dev. Biol.

Facial (nVII) motor neurons are found aberrantly in r4 in trilobite mutants. All panels show dorsal views of the hindbrain with anterior to the left. Double arrows in (B), (D), and (F) denote the midline. (C, D) Composite confocal images of embryos; identify GFP-expressing motor neurons in the fluorescein channel and zn5-labeled commissural neurons and axons in the rhodamine channel. (A) In a 48-HPF wild-type sibling, the islet antibody labels the trigeminal motor neurons (nV) in r2 and r3, the facial motor neurons (nVII) in r6 and r7 (arrows), the glossopharyngeal motor neurons (nIX) in r7 (arrowhead), and the vagal motor neurons (nX) in the caudal hindbrain (chb). The strongly labeled, medially located cells in r5 and r6 surrounding the asterisk are the abducens motor neurons (nVI). (B) In a tri mutant, the nV, nVI, and nX neurons are found in the same locations as in wild-type siblings. A large number of islet-labeled cells are found in r4 (arrow), corresponding to nVII neurons. The nIX neurons in r7 are absent. (C) In a 48-HPF wild-type sibling, the zn5 antibody labels commissural axons at rhombomere boundaries in a characteristic fashion and confirms that the GFP-expressing nVII neurons (arrows) are located in r6 and r7. (D) In a tri mutant, zn5 labeling confirms that nVII neurons (arrow) are located in r4. (E) In a 36-HPF wild-type sibling, application of the lipophilic dye, DiI, to the hyoid arch retrogradely labels nVII cell bodies in r6 and r7 (arrows). (F) By contrast, in a tri mutant, DiI application in the hyoid arch retrogradely labels nVII cell bodies in r4 exclusively (arrow). oto, otocyst. Scale bar, 100 (A–D) and 50 μm (E, F). |
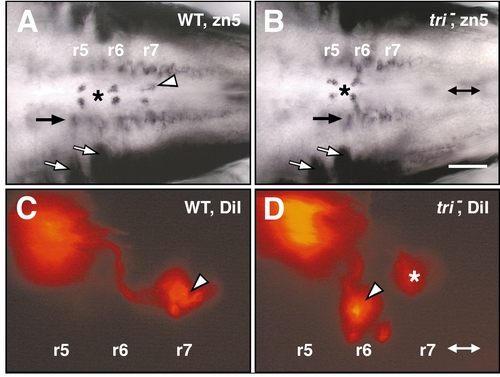
Glossopharyngeal (nIX)-like motor neurons are found in r6 in trilobite mutant embryos. All panels show dorsal views with anterior to the left. Double arrows in (B) and (D) denote the midline. (A) In a 48-HPF wild-type sibling, the zn5 antibody labels abducens (nVI) motor neurons medially in r5 and r6 (surrounding the asterisk) and the nIX motor neurons medially in r7 (arrowhead). (B) In a tri mutant, the nVI motor neurons (asterisk) develop normally in r5 and r6. However, the nIX motor neurons are absent from r7. Other zn5-labeled cell types such as the hindbrain commissural neurons (white arrows) and other uncharacterized cells (black arrow) develop in a similar fashion in wild-type and mutant embryos. (C) In a 48-HPF wild-type sibling, DiI application in the first gill arch retrogradely labels nIX neuron cell bodies in r7 (arrowhead). (D) By contrast, in a tri mutant, DiI application in the first gill arch labels nIX cell bodies exclusively in r6 (arrowhead). The out-of-focus labeled cells in r7 (asterisk) are seen in some wild-type and mutant embryos and appear to be very anteriorly located nX neurons that contain DiI, due to inadvertent labeling of nX axons at the DiI injection site. Scale bar, 100 (A, B) and 50 μm (C, D). |

Facial (nVII) motor neuron migration is eliminated in trilobite mutants. All panels show dorsal views of the hindbrain with anterior to the left. (A–C) In a wild-type sibling (A), putative GFP-expressing nVII neurons accumulate in r4 by 18 HPF. By 24 HPF (B), nVII neurons have started migrating tangentially across rhombomere boundaries into r5, r6, and r7. By 30 HPF (C), most nVII neurons have migrated into r6 and r7. The nV neurons in r2 are induced early and remain in r2 throughout development. The nV neurons in r3 arise after 30 HPF. (D–F) In a tri mutant embryo (D), putative nVII neurons arise in r4, as in wild-type embryos. By 24 HPF (E), the number of GFP-expressing cells in r4 increases, but the cells fail to migrate tangentially (posteriorly). By 30 HPF (F), a large number of nVII neurons have accumulated in r4, and no cells have migrated into r6 and r7. Scale bar, 75 μm. |

Mutant nVII motor axons extend correctly into the hyoid arch. (A) and (B) show dorsal views, and (C) and (D) show side views with anterior to the left in all panels. (A) In a 96-HPF wild-type sibling, most nVII neurons (arrow) are found in r6. (B) In a trilobite mutant, the nVII neurons (arrow) are found exclusively in r4. The few GFP-expressing cells in r6 are not nVII neurons because they do not extend axons into the hyoid arch (n = 16 embryos). (C) In a 48-HPF wild-type sibling, nVII axons (arrow) extend into the hyoid arch. The inset image is a higher magnification view of the boxed area and clearly shows a characteristic branching of the nVII axons. (D) In a tri mutant, the pattern of nVII axon extension into the hyoid arch, including the characteristic branching (inset), is indistinguishable from that of its wild-type sibling. oto, otocyst. |

Hindbrain development and other cell migrations are unaffected in trilobite mutants. (A–F), (I), and (J) show dorsal views, and (G) and (H) show side views, with anterior to the left. (A) In an 18-HPF wild-type sibling, hoxb1a is expressed in r4. (B) In a tri mutant, hoxb1a is expressed in a normal fashion in r4. (C) In a 36-HPF wild-type sibling, the 3A10 antibody labels the Mauthner cell bodies in r4 and their axons. (D) In a tri mutant, the Mauthner cells and their axons develop normally. (E) In a 36-HPF wild-type sibling, gata3 is expressed in nVII motor neurons in r6 and r7 (arrowheads), nV neurons in r2 (asterisk), and in nVIII sensory neurons (out-of-focus cells, arrow). (F) In a tri mutant, the gata3-expressing nVII neurons are located in r4 (arrowhead). (G) In a 36-HPF wild-type sibling, an anti-tyrosine hydroxylase antibody labels locus coeruleus (LC) neurons ventrally within r1. (H) In a tri mutant, the LC neurons are found at the same location as in wild-type embryos. (I) In a wild-type sibling, the F59 antibody labels adaxial cells (arrowhead) adjacent to the notochord at the 18-somite stage. By the 30-somite stage, the F59-labeled cells (arrowhead) have migrated radially and laterally away from the notochord and have differentiated into superficial slow muscle fibers. (J) In a tri mutant, the F59-labeled cells (arrowheads) are shorter due to decreased convergence and extension. The location of labeled cells at 18- and 30-somite stages is identical to that in wild-type embryos, indicating that adaxial cell migration occurs normally. oto, otocyst. Scale bar, 100 (A, B, G, H, “30 som” I, J) and 50 μm (C–F, “18 som” I, J). |

nVII motor neurons migrate normally in the gastrulation mutant, knypek (kny). All panels show dorsal views of the islet antibody-labeled hindbrains with anterior to the left. Embryos were from obtained from a cross of trilobite;knypek double heterozygotes. The arrowheads in (B–D) point to the most anterior spinal motor neurons. The strongly labeled, medially located cells in r5 and r6 surrounding the asterisk are the abducens (nVI) motor neurons. (A) In a 36-HPF wild-type sibling, the islet-labeled cells corresponding to the nV, nVII (arrows), and nX neurons are found at their characteristic locations. (B) In a knypek mutant, the islet-labeled cells corresponding to the nV, nVII (arrows), and nX motor neurons are found at their normal locations, as in wild-type embryos. (C) In a trilobite mutant, the nVII neurons (arrow) are found in r4. (D) In a trilobite;knypek double mutant, islet-labeled cells corresponding to putative nVII neurons (arrow) are still located in r4. The nVI neurons in r6 are missing. The nX neurons are greatly reduced in number and are positioned very laterally compared to either single mutant, reflecting the extremely reduced convergence extension cell movements in double-mutant embryos. Scale bar, 100 μm. |
Reprinted from Developmental Biology, 242(2), Bingham, S., Higashijima, S.-I., Okamoto, H., and Chandrasekhar, A., The zebrafish trilobite gene is essential for tangential migration of branchiomotor neurons, 149-160, Copyright (2002) with permission from Elsevier. Full text @ Dev. Biol.